Class
Classification
Subordinate Taxa
Nomenclature
Scientific Name:
Lembophyllaceae Broth.
Type Taxon:
Description
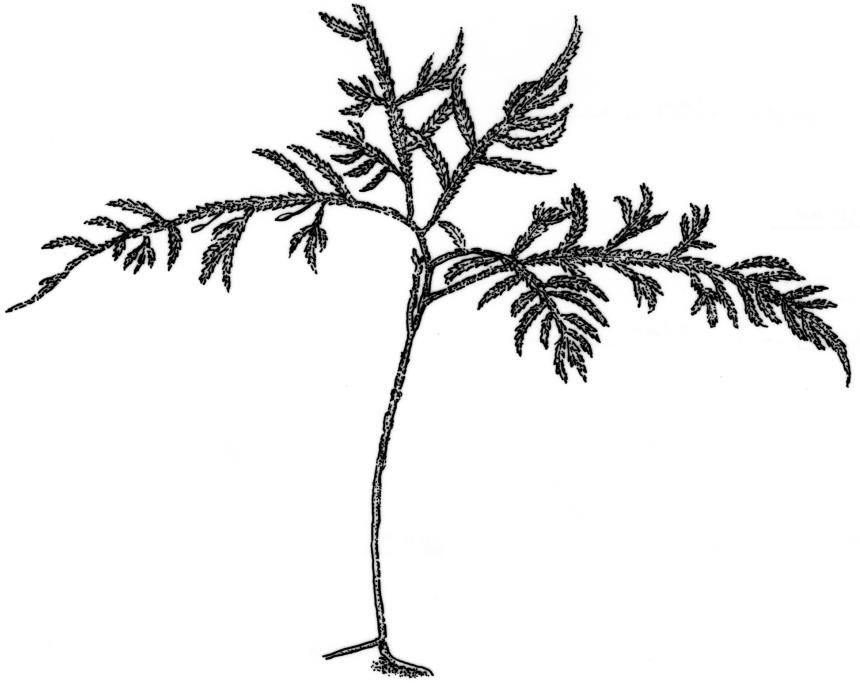
Plants robust to slender, rigid and wiry or soft, often glossy. Stems mostly erect-ascendant, sometimes pendent or trailing, subdendroid to dendroid and stipitate, loosely or densely branched to form usually regular fronds, terete to complanate; paraphyllia absent. Pseudoparaphyllia foliose. Leaves erect-spreading, rarely patent or falcate-secund, moderately to deeply concave, smooth or ± wrinkled when dry; ovate to widely ovate or ± round, apices obtuse or widely acute, rarely cucullate, acute, acuminate, cuspidate or apiculate, rarely aciculate; margins plane, incurved, unbordered, entire or variably denticulate. Costa variable, mostly double and short, occasionally single and developed to mid leaf, faint, sometimes absent. Mid laminal cells linear or linear-rhombic, prorate and sometimes porose; becoming shorter and rhombic above; cells near base becoming shorter, wider, more irregular and porose, sometimes forming a basal band; alar cells short, quadrate, sometimes not distinct from adjacent basal cells.
Dioicous or pseudautoicous. Inner perichaetial leaves mostly widely spreading from an oblong base, sometimes erect, mostly acute to acuminate or lanceolate, ecostate, not or weakly bordered; areolation as in stem leaves, porose and prorate, alar cells not differentiated, margins entire, enclosing numerous filamentous hyaline paraphyses. Outer perichaetial leaves shorter and more obtuse. Perigonia axillary on lateral branches, gemmiform, with leaves enclosing three to several antheridia with hyaline paraphyses. Setae elongate, red-brown, curved or flexuose, smooth. Capsules mostly horizontal or sometimes ± erect, symmetric, smooth, often constricted below the mouth when dry; exothecial cells mostly irregularly thick-walled, elongate, not or sometimes weakly thickened at the angles. Stomata usually absent. Peristome double, hypnoid; exostome teeth yellow-brown, lanceolate, shouldered, bordered below, inserted at or below the capsule mouth, not furrowed; endostome pale yellow or hyaline, papillose, with a basal membrane c. (⅕–) ⅓–½ the exostome teeth in length, segments and cilia mostly well developed. Operculum conic, blunt-apiculate or rostrate. Calyptra narrowly cylindric, cucullate, smooth and obtuse, hairless or rarely with 2–3 hairs at the base. Spores spherical, light yellow-brown, papillose.
Taxonomy
The Lembophyllaceae are a moderately sized and widely distributed temperate and tropical family of pleurocarpous mosses. The generic limits of the family have been unstable. Brotherus (1925) originally included 12 genera in the Lembophyllaceae, but as subsequent authors questioned many of those placements, the family came to be viewed more narrowly, containing as few as two species (Buck & Vitt 1986; Crum 1991; Buck 1994). More recently, molecular analysis supports the recognition of a family concept close to that of Brotherus, comprising 14 genera. Perceived relationships of the family have also shifted, away from families such as the Brachytheciaceae and the Meteoriaceae, to a close relation to the Neckeraceae (Tangney 1997; Quandt et al. 2009; Enroth et al. 2019).
The genera included in the Lembophyllaceae exhibit a diversity of peristomes, having both well-developed and reduced forms, with the latter associated with an epiphytic habit (Buck 1991; Quandt & Huttunen 2004; Huttunen et al. 2004). Within the family, epiphytic members of the main clades exhibit peristome reduction alongside taxa with well-developed peristomes. Weymouthia mollis is the only N.Z. Lembophyllaceae with a reduced peristome.
The Lembophyllaceae comprise three main clades/groups broadly distributed in the temperate Northern Hemisphere, South America – Africa, and Australasia – Southern South America respectively (Tangney 2007a).
Five genera, Lembophyllum, Weymouthia, Camptochaete, Fifea and Fallaciella, with 11 species, including one variety, are accepted as part of the New Zealand flora.
Quandt et al. (2009), using chloroplast markers trnL-trnF and pbsT-psbH and ITS2, showed that these five genera belong in Lembophyllaceae but sequence divergence is very low between the genera and their study could not test the monophyly of Fifea and Fallaciella.
Key
| 1 | Plants mostly robust, or rigid and wiry, forming scrambling wefts, occasionally pendent, or dendroid and stipitate; secondary stems differentiated into a lower unbranched stipe with erect-appressed leaves and upper much-branched fronds with erect-spreading leaves | 2 |
| 1' | Plants robust or slender, mostly forming soft, tight mats, loose wefts or pendent masses, not dendroid or stipitate; secondary stems with leaves not erect-appressed below, or stipe poorly developed, not frondose, sparsely branched | 4 |
| 2 | Frond axis and branch leaves aciculate, acumen greater in length than the rest of the leaf; apical cell linear, longer than 75 µm | Fifea |
| 2' | Frond axis and branch leaves not aciculate, apex obtuse or acute to acuminate; apical cell rhombic to short linear, less than 50 µm | 3 |
| 3 | Frond axis and branch leaves various, ovate to widely ovate or cochleariform; apices obtuse or acute to acuminate; costa double and short; mid laminal cells linear | Camptochaete |
| 3' | Frond axis and branch leaves not various, cochleariform; apices obtuse, cucullate, less commonly widely acute; costa variable; mid laminal cells rhombic to short linear | Lembophyllum |
| 4 | Frond axis and branch leaves aciculate, acumen greater in length than the leaf base; apical cell linear, longer than 75 µm | Fifea |
| 4' | Frond axis and branch leaves not aciculate, apex acute to acuminate; apical cell rhombic to short linear, less than 50 µm | 5 |
| 5 | Plants small, forming tight or rather loose mats; leaves moderately concave, ovate, broadly to narrowly acute; apex erect, not cucullate, sometimes mucronate-apiculate | Fallaciella |
| 5' | Plants large, forming loose mats or wefts, sometimes pendent; leaves concave to deeply concave, cochleariform, to widely ovate or ovate oblong; apex cucullate to widely acute, not mucronate-apiculate | 6 |
| 6 | Plants usually weft-forming, stems julaceous, leaves cochleariform; apex usually cucullate; costa varying from short and double or absent to robust and single; mid laminal cells rhombic to short linear, thick-walled, sometimes weakly porose | Lembophyllum |
| 6' | Plants usually or at least sometimes pendent, stems complanate, sometimes loosely julaceous, leaves ovate to oblong; apex erect; costa short and double or absent; mid laminal cells linear, firm-walled, sometimes strongly porose | Weymouthia |
Recognition
The N.Z. members of the family share a common branching pattern that produces a range of growth forms, from relatively coarse wiry wefts to low smooth mats or soft pendent fronds. They are generally ± glossy plants with ovate-oblong, concave leaves, and a distinct cell pattern of rhombic upper laminal cells, linear mid laminal cells which are usually prorate, and differentiated basal and alar cells. The costa varies from short and double to faint or absent, and is occasionally single and extending to mid leaf. Capsules are mostly exserted on elongated setae with generally well-developed hypnoid peristomes.
In mesic and xerophytic habitats, they may be confused with members of the Brachytheciaceae, but the latter tend to have a strong single costa and typically have linear upper laminal cells. The Brachytheciaceae also lack the wiry stipe-forming stems of the Lembophyllaceae. Similarly, the Amblystegiaceae are unlikely to be confused. In wetter terrestrial habitats favoured by many Amblystegiaceae, species of Lembophyllaceae are not common. The Amblystegiaceae are often red-brown, yellow-green or sometimes blackish and have leaves that are often strongly curved to one side, with a costa single to above mid leaf, mostly elongate upper laminal cells, and alar cells that are often thin-walled and inflated. Occurrence in wetter habitats for the Lembophyllaceae is mostly in stream beds, on rocks, bark and soil, and there they are most similar to members of the Neckeraceae, particularly the dendroid-forming Austrothamnium pandum and Echinodiopsis hispida. The latter two are distinguished by strong single costae, serrate leaf margins, and laminal cells that are densely packed and irregularly quadrate to round. Other families that produce creeping stems with erect dendroid fronds are similarly distinct from the Lembophyllaceae. These include the umbrella mosses of the Hypopterygiaceae (which have flattened fronds with usually bordered and dentate leaves in three rows; two rows of larger asymmetric leaves and a third row of smaller, symmetric under-leaves, with strong single costae), and the Hypnodendraceae (that have branches in whorls rather than pinnate fronds, except Braithwaitea sulcata, which has leaves with strong single costae, to the leaf apex or excurrent, and laminal cells narrowly linear above). Trachyloma, in the Pterobryaceae produces flattened pinnate fronds on unbranched stipes, with leaves that are ovate to ovate-lanceolate and have a costa that is ± absent. However, it differs from the Lembophyllaceae in having a silver-white colouring, cells narrowly elongate mostly throughout, becoming laxer at the base, and numerous brown brood bodies amongst the apical branch leaves. The pendulous forms of Weymouthia with distantly and irregularly pinnate branches are similar to those of Papillaria in the Meteoriaceae, but Papillaria tends to be more strikingly coloured, with shades of orange, yellow, brown and yellow-green. The leaves are often plicate, with a strong single costa extending to the upper leaf, they lack differentiated alar cells, and the plants are dull, due to the leaf cells being strongly and multiply papillose.
Biostatus
Indigenous (Non-endemic)
| Category | Number |
|---|---|
| Indigenous (Endemic) | 3 |
| Indigenous (Non-endemic) | 8 |
| Total | 11 |
Notes
A common, dendroid-stipitate, branching pattern underlies the range of form in N.Z. Lembophyllaceae. Plants produce branched fronds on unbranched stems (stipes) forming small ‘trees’ linked together by creeping stems. Fronds develop as erect side branches from creeping stems, and by the creeping stems becoming erect to form fronds. New growth may occur from the frond branches, as they grow downwards and root at the tips to form new creeping stems. Creeping stems and the unbranched stipes are characterised by leaves which are pressed to the stems making them appear leafless. Changes from one branch type to another can be traced through changes in orientation of the leaves (Tangney 2007b).
The N.Z. genera exhibit variations on the underlying pattern of growth and in the degree of frond formation. For example, Camptochaete forms mostly pinnate fronds with wiry stems, whereas in Weymouthia the fronds are only loosely branched, or fronds are lacking, and softer pendent growth is typical. These reductions in the branching are associated with an epiphytic habit (Tangney 2007b).
Variability of this pattern of growth and branching often produces elongate creeping or pendent forms where the separation of creeping stems and fronds is not clear as the orderly production of typical growth breaks down. Such forms are characteristic of some of the N.Z. species, making them difficult to identify as the distinctions between species are blurred.
Extensive tangled masses are sometimes produced as loose-lying, unattached ‘moss balls’ (Martin 1952). Martin noted this tendency in plants of Camptochaete (ramulosa) deflexa, and they are extreme forms of the elongate forms and are sometimes found in wet depressions under scrub and forest, where ongoing disturbance seems to be a factor in continually producing regenerative growth (Tangney 2007b).
A number of the species are pseudautoicous, with dwarf males reported in Lembophyllum (Tangney 2006) and in Camptochaete, Fifea, and Weymouthia by Hedenäs & Bisang (2011).
Excluded Taxa
Acrocladium chlamydophyllum (Hook.f. & Wilson) Müll.Hal. & Broth. Brotherus (1901–1909, 1925) included Acrocladium in the Lembophyllaceae, and subsequent authors have either excluded it (Buck & Vitt 1986, Buck 1991, Walther 1983; Tangney 1997) or included it (Crosby et al. 1999). Molecular data supports its exclusion from the Lembophyllaceae, and it has been placed in a monogeneric family, the Acrocladiaceae, with relations to the Lepyrodontaceae (Tangney et al. 2010; Quandt et al. 2009; see also, Fife 2014).
Bibliography
Brotherus, V.F. 1901–1909: Musci (Laubmoose) II Specieller Teil. In: Engler, A.; Prantl, K. (ed.) Die natürlichen Pflanzenfamilien. Teil 1. Abt. 3. Engelmann, Leipzig. 277–1246.
Brotherus, V.F. 1925: Musci (Laubmoose). In: Engler, A. (ed.) Die natürlichen Pflanzenfamilien. Edition 2. Bd 11. Engelmann, Leipzig. 1–542.
Buck, W.R. 1991: The basis for familial classification of pleurocarpous mosses. Advances in Bryology 4: 169–185.
Buck, W.R. 1994: A new attempt at understanding the Meteoriaceae. Journal of the Hattori Botanical Laboratory 75: 51–72.
Buck, W.R.; Goffinet, B. 2000: Morphology and classification of mosses. In: Shaw, A.J.; Goffinet, B. (ed.) Bryophyte Biology. Cambridge University Press, Cambridge. 71–123.
Buck, W.R.; Vitt, D.H. 1986: Suggestions for a new familial classification of pleurocarpous mosses. Taxon 35(1): 21–60.
Crosby, M.R.; Magill, R.E.; Allen, B.; He, S. 1999: A Checklist of the Mosses. Missouri Botanical Garden, St. Louis.
Crum, H.A. 1991: A partial clarification of the Lembophyllaceae. Journal of the Hattori Botanical Laboratory 69: 313–322.
Enroth, J.; Olsson, S.; Huttunen, S.; Buchbender, V.; Tangney, R.; Stech, M.; Hedenäs, L.; Quandt, D. 2019: Orthostichellaceae fam. nov. and other novelties in pleurocarpous mosses revealed by phylogenetic analyses. Bryologist 122(2): 219–245. (Published online: 20 May 2019)
Fife, A.J. 2014: Amblystegiaceae. In: Heenan, P.B.; Breitwieser, I.; Wilton, A.D. (ed.) Flora of New Zealand — Mosses. Fascicle 1. Manaaki Whenua Press, Lincoln.
Goffinet, B.; Buck, W.R.; Shaw, A.J. 2009: Morphology, anatomy, and classification of the Bryophyta. In: Goffinet, B.; Shaw, A.J. (ed.) Bryophyte Biology. Edition 2. Cambridge University Press, Cambridge. 55–138.
Hedenäs, L.; Bisang, I. 2011: The overlooked dwarf males in mosses—unique among green land plants. Perspectives in Plant Ecology, Evolution and Systematics 13(2): 121–135.
Huttunen, S.; Ignatov, M.S.; Muller, K.; Quandt, D. 2004: Phylogeny and evolution of epiphytism in three moss families Meteoriaceae, Brachytheciaceae, and Lembophyllaceae. Monographs in Systematic Botany from the Missouri Botanical Garden 98: 328–361.
Martin, W. 1952: Unattached mosses in the New Zealand flora. Bryologist 55: 65–79.
Quandt, D.; Huttunen, S. 2004: Evolution of pendent life-forms in bryophytes. Journal of the Hattori Botanical Laboratory 95: 207–217.
Quandt, D.; Huttunen, S.; Tangney, R.; Stech, M. 2009: Back to the future? Molecules take us back to the 1925 classification of the Lembophyllaceae (Bryopsida). Systematic Botany 34(3): 443–454.
Tangney, R.S. 1997: A generic revision of the Lembophyllaceae. Journal of the Hattori Botanical Laboratory 81: 123–153.
Tangney, R.S. 2006: Phyllodioicy in Lembophyllum (Bryopsida: Lembophyllaceae). Journal of Bryology 28(4): 382–384.
Tangney, R.S. 2007a: Biogeography of austral pleurocarpous mosses: distribution patterns in the Australasian region. In: Newton, A.E.; Tangney, R.S. (ed.) Pleurocarpous Mosses, Systematics and Evolution. CRC Press, Boca Raton. 393–407.
Tangney, R.S. 2007b: Sympodial and monopodial growth in mosses: examples from the Lembophyllaceae (Bryopsida). In: Newton, A.E.; Tangney, R.S. (ed.) Pleurocarpous Mosses, Systematics and Evolution. CRC Press, Boca Raton. 309–319.
Tangney, R.S. 2026: Lembophyllaceae. In: Glenny, D. (ed.) Flora of New Zealand — Mosses. Fascicle 51. Manaaki Whenua Press, Lincoln.
Tangney, R.S.; Huttunen, S.; Stech, M.; Quandt, D. 2010: A review of the systematic position of the pleurocarpous moss genus Acrocladium Mitten. Tropical Bryology 31: 164–170.
Walther, K. 1983: Syllabus der Pflanzenfamilien. Kapitel V, 2. Bryophytina. Laubmoose. Gebruder Borntraeger, Berlin.