Class
Family
Subfamily
Genus
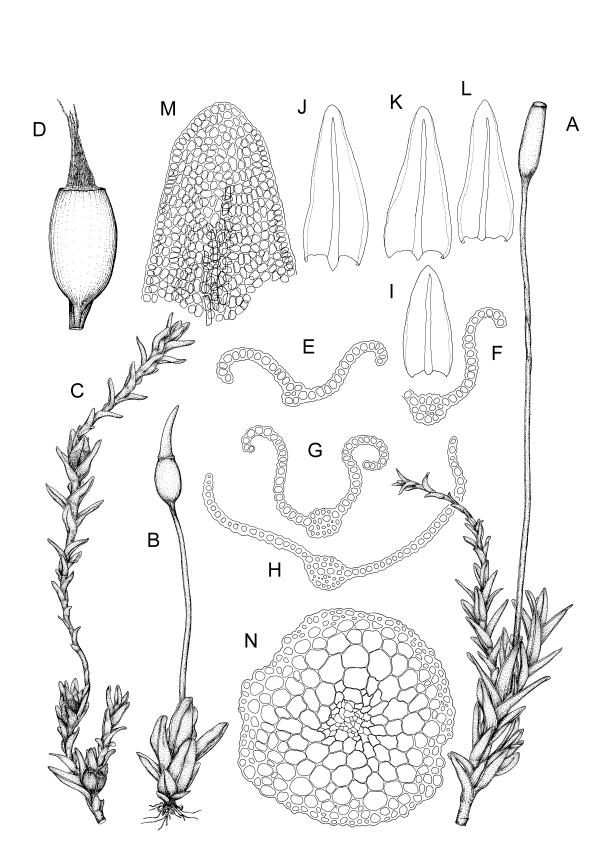
Plants green to dark green or brown, densely or loosely gregarious. Stems ≤ 2(–8) mm, occasionally branched, in cross-section with a central strand and a weakly developed sclerodermis. Leaves erect-spreading when moist, erect and little altered or weakly shrivelled when dry, 0.8–1.6 mm, lingulate, carinate above and often loosely once plicate on one or both sides, rounded and ± flat at apex; margins variably recurved at mid leaf, unistratose, entire or papillose; upper laminal cells clear in outline, irregularly isodiametric, firm-walled, with 1–3 low papillae or apparently smooth, (8–)9–12 × 9–12 µm, becoming oblate towards the margins and longer towards the leaf base; lower laminal cells differentiated, lax, hyaline, rectangular, thin-walled, smooth, with several rows ± quadrate at the margins. Costa ± concolorous, failing c. 8–12 cells before the leaf apex; adaxial superficial cells elongate in at least the distal ½; abaxial superficial cells quadrate to short-rectangular distally. Rhizoidal tubers present, comprising a chain of uniseriate cells c. 20 µm wide, or ovoid, of 8 × 6 cells to c. 135 µm in longest diameter, with cells slightly bulging. Axillary hairs of 2–3 cells of which basal cell is shorter and brown. Laminal KOH colour reaction yellow-orange.
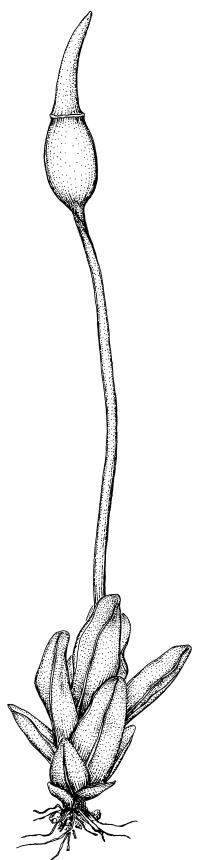
Dioicous. Perichaetia with leaves similar in size and shape to vegetative leaves, but with basal zone of hyaline, rectangular, thin-walled, smooth cells extending to c. ½ leaf length in innermost perichaetial leaf. Perigonia terminal, becoming lateral by innovation, bulbiform. Setae slender, flexuose, red at base, paler above, 3–6(–8) mm. Capsules erect, short- to long-ellipsoid, 0.5–1.7 mm. Operculum rostrate, with a slightly curved beak, ± equal the theca, with cells in straight rows. Peristome of 16 teeth, 300–500 µm, pale buff or colourless, often persistent in older capsules, each tooth divided ± completely into 2 slender rami, densely and finely spiculose throughout and scarcely articulated, straight, arising from a low basal membrane, which is similarly ornamented. Spores 7.5–11(–13) µm, smooth.
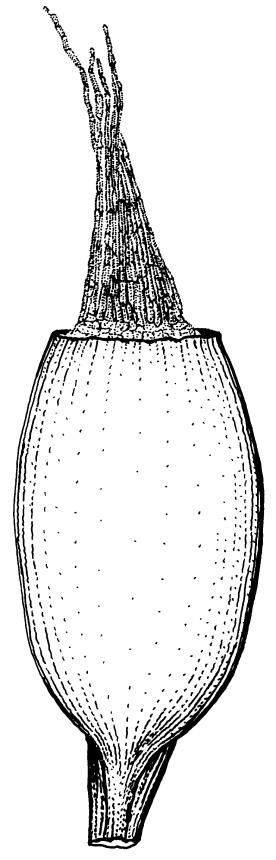
Wilson 1854, pl. 84, fig. 4 (as Trichostomum lingulatum); Lüth 2019, p. 633 (as Didymodon tophaceus); Malcolm et al. 2020, pp. 524–525 (as Didymodon tophaceus)
The typical form of Geheebia tophacea in N.Z. is well described by Sainsbury (1955) (as Desmatodon lingulatus) in having plants "minute … consisting of a brown or brown-green bulbil of appressed leaves" (Beever 2024, fig. B), and the leaves being "widely oblong-lingulate". Confusion is most likely with allied species that are small in stature, and like G. tophacea have rounded-obtuse leaf apices with the costa failing before the apex. These are G. ceratodontea (for distinctions refer to that species), and three diminutive species of Tridontium: T. cockaynei, T. milleneri, and T. novae-zelandiae (for distinctions refer to the key to the genera of subfamily Barbuloideae).
K; NI: N Auckland, S Auckland, Gisborne, Hawke’s Bay, Wellington; SI: Marlborough (D’Urville I.), Canterbury, Otago, Southland.
Recorded from Campbell I. by Crum & Anderson (1981) but no specimen has been seen for the present study.
Widespread in both hemispheres. Recorded from mainland Australia by Streimann & Klazenga (2002).
On soil or soft rock, typically calcareous, and often damp. "Papa" is the most frequently recorded substrate, but "midden deposits" and "volcanic tuff" are also noted on herbarium specimens.
Mixed gatherings with related species Trichostomopsis australasiae, Geheebia ceratodontea, and Gertrudiella torquata are frequent. Other associated mosses are species of Bryum, including B. sauteri, but also Fissidens leptocladus and Funaria hygrometrica.
Geheebia tophacea is typically a lowland moss in N.Z., with several records close to sea level, and most below 400 m elevation. The highest known specimen (Ohakune, R. Mundy 155, CHR 570269) is from an elevation of c. 600 m.
The decision by Zander (1993 p. 341) to place Trichostomum lingulatum Hook.f. & Wils. in synonymy with the subcosmopolitan Geheebia tophacea (as Didymodon tophaceus) is accepted here. The N.Z. moss shares the rounded leaf apex, failing costa, and long, straight, irregularly cleft peristome teeth of European material, and all these features are well illustrated in the protologue of Trichostomum lingulatum (Wilson 1854, Tab LXXXIV, fig. 4). In addition, both taxa have elongate cells on the adaxial face of the costa in its distal half, a feature regarded as characteristic of G. tophacea by Jiménez (2006) and Zander et al. (2007) (both as Didymodon tophaceus). Zander et al. (2007) suggest that "insufficient evidence has been placed on the elongate cells of the adaxial costal surface of Didymodon tophaceus".
Rhizoidal tubers of the form illustrated by Arts & Sollman (1998) for an Ecuadorian specimen (as D. tophaceus) have been observed in N.Z. material from Invercargill, Southland L.D. (D. Grantham s.n., CHR 570624), and Aorangi Range, Wellington L.D. (J.E. Beever 94-17a, WELT M044317).
In the wider distribution of Geheebia tophacea, leaves are described as having a broadly ovate base that is often, but not invariably, decurrent on the stem (e.g., see treatments for Britain and Ireland (Smith 2004), North Africa, Europe, and Asia (Jiménez 2006), Zander et al. (2007) for North America, all as Didymodon tophaceus). In N.Z. material leaves are lingulate and are only rarely decurrent on the stem; e.g., in a specimen from Waihua, Hawke’s Bay L.D. (G.O.K. Sainsbury 16102, WELT M014284).
The multi-locus phylogeny published by Jiménez et al. (2022) did not include any southern hemisphere samples of this species, and attempts to obtain DNA sequences from N.Z. specimens of G. ceratodontea were unsuccessful (J. Jiménez, pers. comm., 13 Sep. 2021). Future study may clarify both the relationship of N.Z. G. tophacea with the species elsewhere in its range and its relationship to G. ceratodontea.
Examination of material in Herbarium Hookerianum labelled "Trichostomum lingulatum var." (Colenso s.n., BM 000845867) indicates that the variety β does not warrant recognition.