Class
Family
Subfamily
Genus
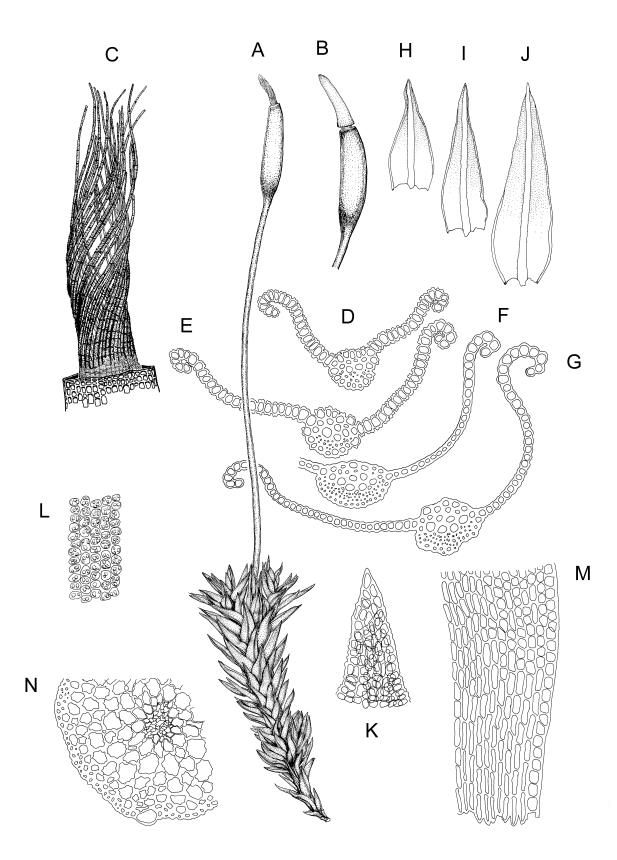
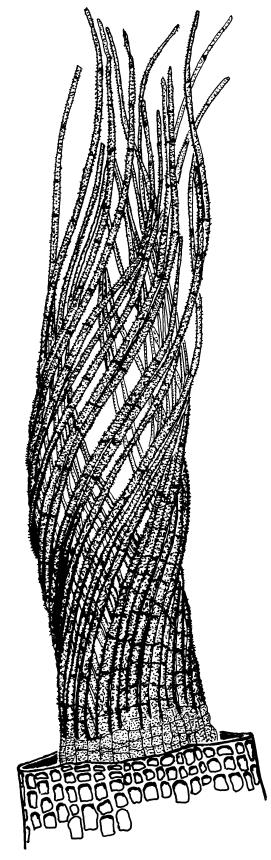
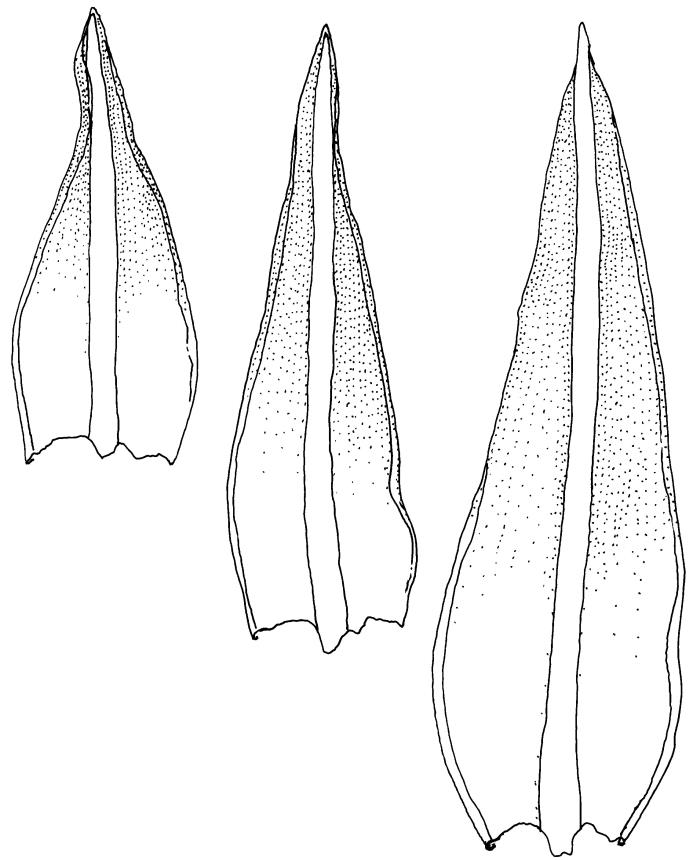
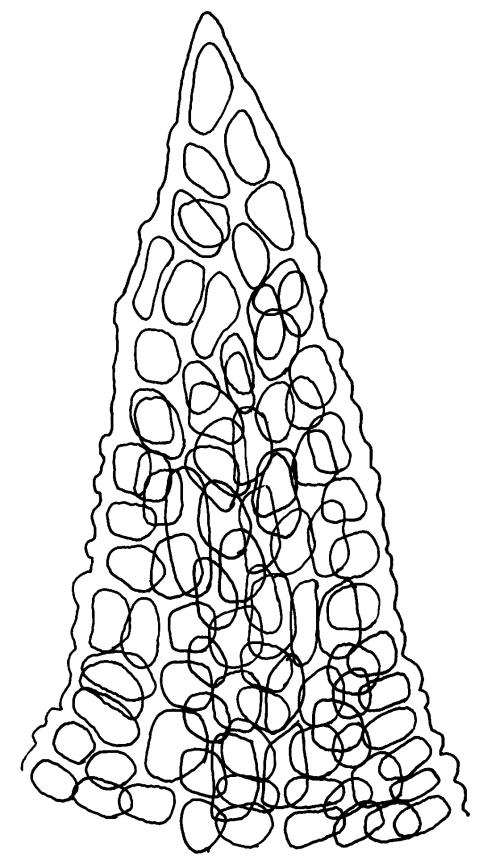
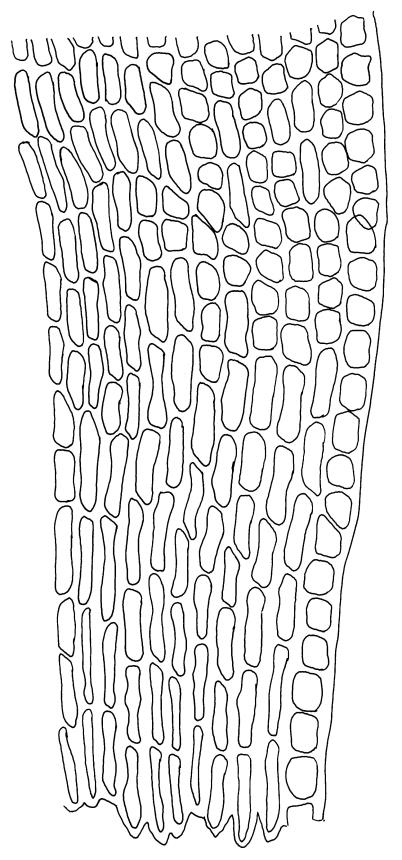
Plants yellow-brown or yellow-green above, dull brown below, in dense tufts or turves, or gregarious. Stems 5–25(–35) mm, branched, in cross-section with a central strand and well-developed sclerodermis. Leaves erect-spreading from a weakly sheathing base when moist, individually twisted and stiffly twisted around the stem to form a rope-like shoot, but often with some apices spreading (especially at the shoot apex) when dry, (1.0–)1.5–3.0 mm, lanceolate, carinate, narrowly obtuse, acute or acuminate; margins narrowly recurved to revolute, unistratose, crenulate by bulging cells and low, rounded papillae; upper laminal cells clear in outline, irregularly rounded-quadrate, thick-walled, bulging, weakly to conspicuously papillose with 1–2 simple or complex papillae, c. 6–8 × 6–8 µm, becoming oblate towards the margin and longer towards the leaf base; lower laminal cells often little differentiated, firm-walled, not hyaline, smooth, rectangular medially, becoming shorter towards the leaf margins, where 1 or more rows are ± quadrate. Costa reddish or concolorous, stout, percurrent or excurrent as a sharp or blunt arista; adaxial superficial cells similar to adjacent laminal cells throughout; abaxial superficial cells quadrate to short-rectangular distally. Axillary hairs of 3–5 cells, with the basal cell shorter and brown. Laminal KOH colour reaction yellow-orange.
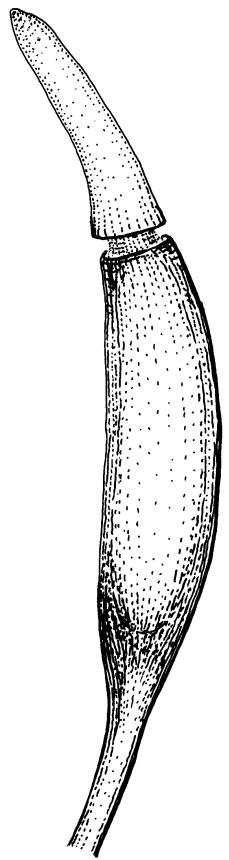
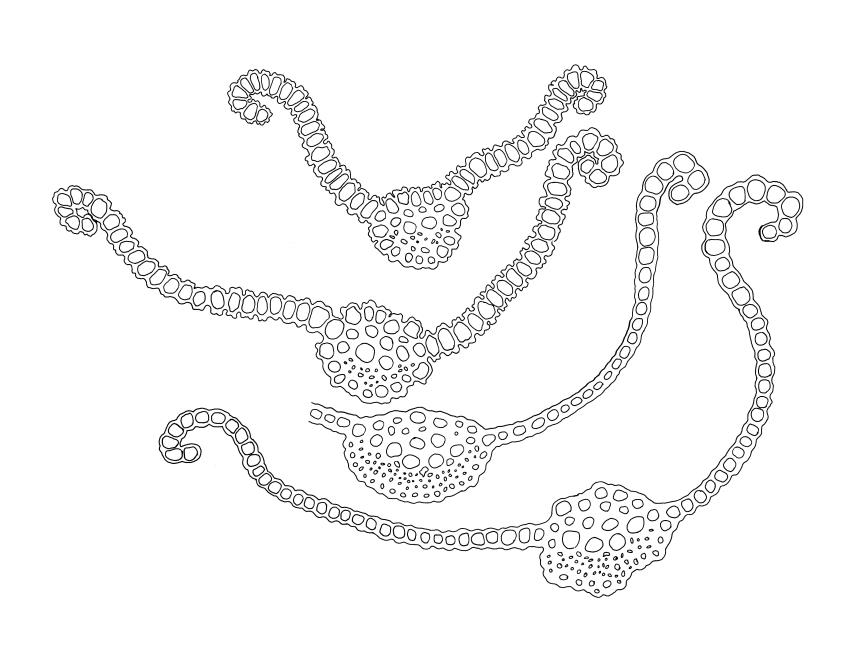
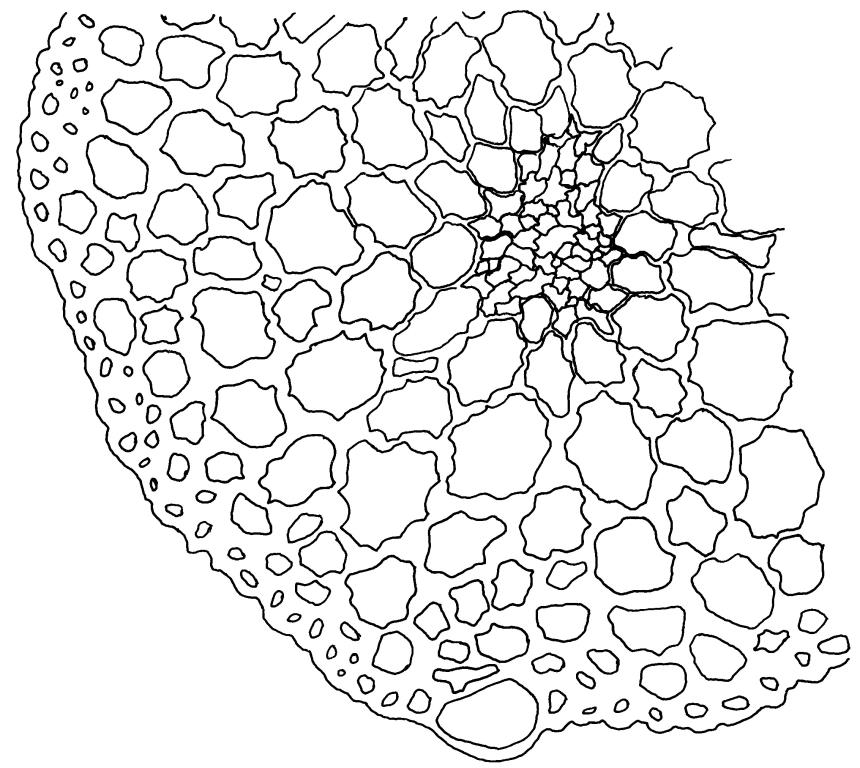
Dioicous. Perichaetia terminal, with perichaetial leaves erect and loosely sheathing the seta base, similar in size and shape to vegetative leaves, or with a longer and more acuminate apex, and with a basal zone of lax, elongate cells, which may extend for the entire lamina in the innermost perichaetial leaf. Perigonia terminal (with innovative shoot produced from below), bulbiform. Setae slender, flexuose, dark red below, paler above, 9–20 mm. Capsules erect, narrowly cylindric and slightly curved, 1.75–2.0 mm. Operculum bluntly and obliquely rostrate, with cells spiralled, c. ½ the theca length. Peristome long (c. 1 mm at dehiscence), spiralled approximately twice, of 16 pale teeth each divided into 2 densely spiculose filiform rami, arising from a low, fenestrate basal cylinder. Calyptra cucullate. Spores 6–10 µm, smooth.
Scott & Stone 1976, pl. 38 (as Barbula torquata); Catcheside 1980, figs 86, 87 (as Didymodon torquatus); Malcolm et al. 2020, pp. 526–528 (as Didymodon torquatus).
The illustration of the dry shoot of Australian material (Scott & Stone 1976) shows a more irregular leaf arrangement than the neat, rope-like form typical in N.Z.
Gertrudiella torquata is a variable moss, but can usually be recognised by its lanceolate leaves, somewhat individually twisted and stiffly twisted round the stem when dry, to form a neat, rope-like shoot, but often with some leaf apices spreading, especially at the shoot apex. The leaf margins narrowly recurved from near leaf base to apex, and the long twisted peristome under an operculum with cells spirally arranged, are further distinguishing features. The form of the leaf apex is confusingly variable. One form is well described by Scott & Stone (1976): "the nerve is thick, excurrent in a rather broad flattish short point."
Confusion with Barbula unguiculata can occur, as it also has neat, rope-like dry shoots and leaves with excurrent costae. In Gertrudiella torquata, however, when the costa is excurrent it extends straight from an acuminate lamina apex and is covered adaxially in its distal part with cells similar to adjacent laminal cells (i.e., quadrate, papillose, and chlorophyllose). In contrast, the costa in B. unguiculata extends from a broadly acute or obtuse lamina apex, and is occasionally recurved. The superficial cells are elongate on both faces, and can be seen with a hand lens as a contrasting pale line in a groove on the adaxial face of the leaf.
Gertrudiella torquata is sometimes confused with Ceratodon purpureus, but its entire leaf margins and papillose laminal cells (vs the presence of some toothing at the apex of at least some leaves, and the smooth laminal cells of C. purpureus) will distinguish them.
K; NI: N Auckland, including offshore islands (GB, RT), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; Ch; A.
Austral. Australia*, Chile*.
A common species on a wide range of soil and rock types, especially calcareous, and including concrete, mortar, shell paths, and midden material, usually in relatively dry, insolated sites. Commonly associated mosses include species of Barbula, Ceratodon purpureus, Tortella flavovirens (in coastal sites), Tortula muralis, and Trichostomopsis australasiae.
Typically a lowland moss, with most records from between sea level and 100 m elevation. G. torquata does, however, extend to high elevations; e.g., at 1400 m in the Arthur Range, Nelson L.D. (A.J. Fife 7498, CHR 438879).
Tortula crispifolia was noted by Mitten (Hooker 1867, p. 419) to differ from Gertrudiella torquata (as Tortula torquata) by the costa being "excurrent into a pungent mucro", but to be "scarcely excurrent" in the latter. After examining both types, the present author agrees with Dixon (1923, pp. 128–129) that the two should be synonymised.
Gertrudiella torquata (as Didymodon torquatus) was placed in synonymy of the morphologically similar Didymodon vinealis (Brid.) R.H.Zander by Sollman (1996). The latter species was subsequently transferred (Zander 2013) to a new genus, Vinealobryum R.H.Zander. On the basis of DNA sequencing, Jiménez et al. (2022) demonstrated that the two species are only distantly related, and justifiably are placed in separate genera.
The molecular data that supports the transfer of Didymodon torquatus to Gertrudiella (Jiménez et al. 2022) comprised sequences obtained from two N.Z. Hawke’s Bay specimens (roadside bank, vicinity of Dannevirke, J.E. Beever 109-66, CHR 629129; bank of Tukituki River, J.E. Beever 109-80b, CHR 629130). These two specimens nest deeply among 14 other taxa, including three South American specimens of the type taxon of the genus, G. validinervis (Herzog) Broth. (as G. uncinicoma (Müll.Hal.) G.M.Suárez & Schiavone.