Class
Family
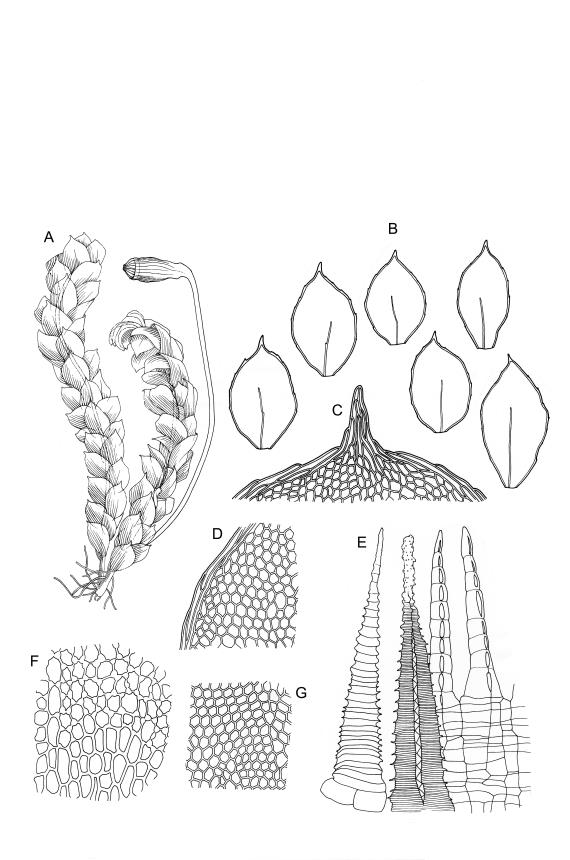
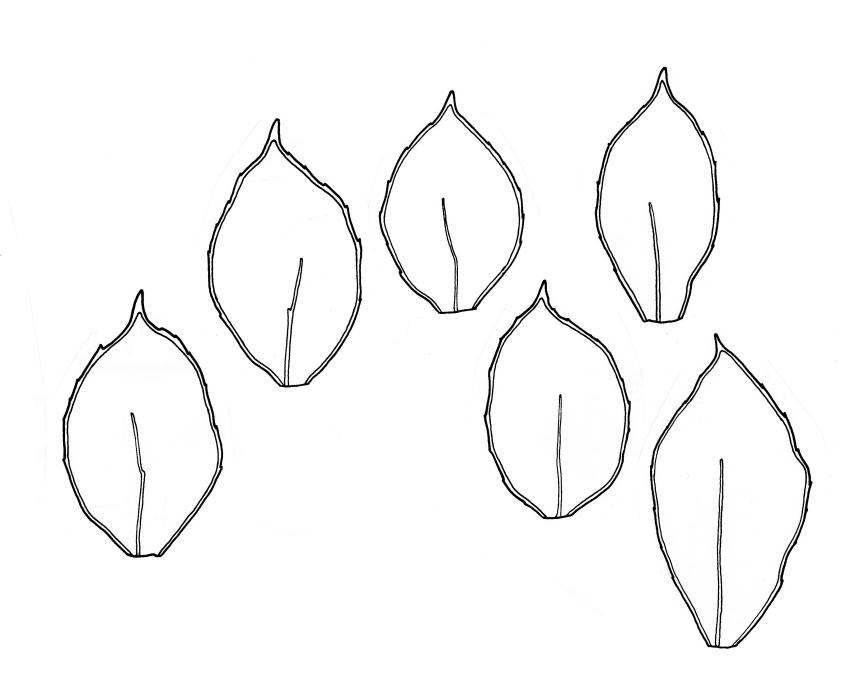
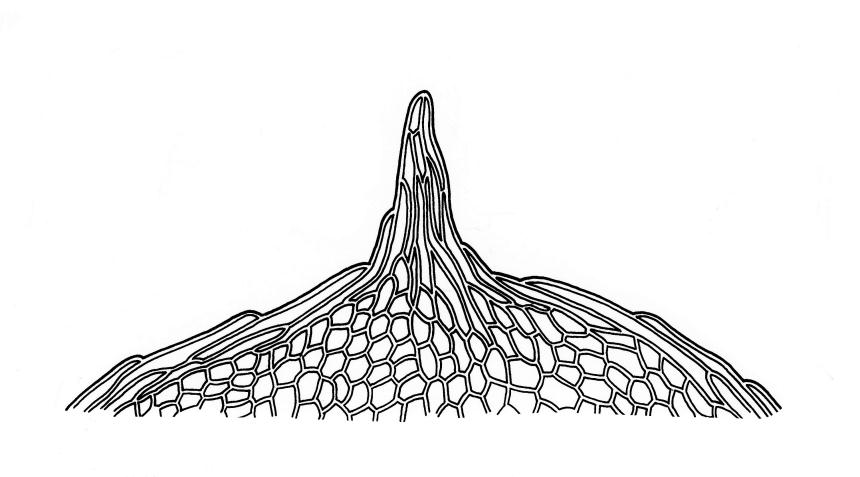
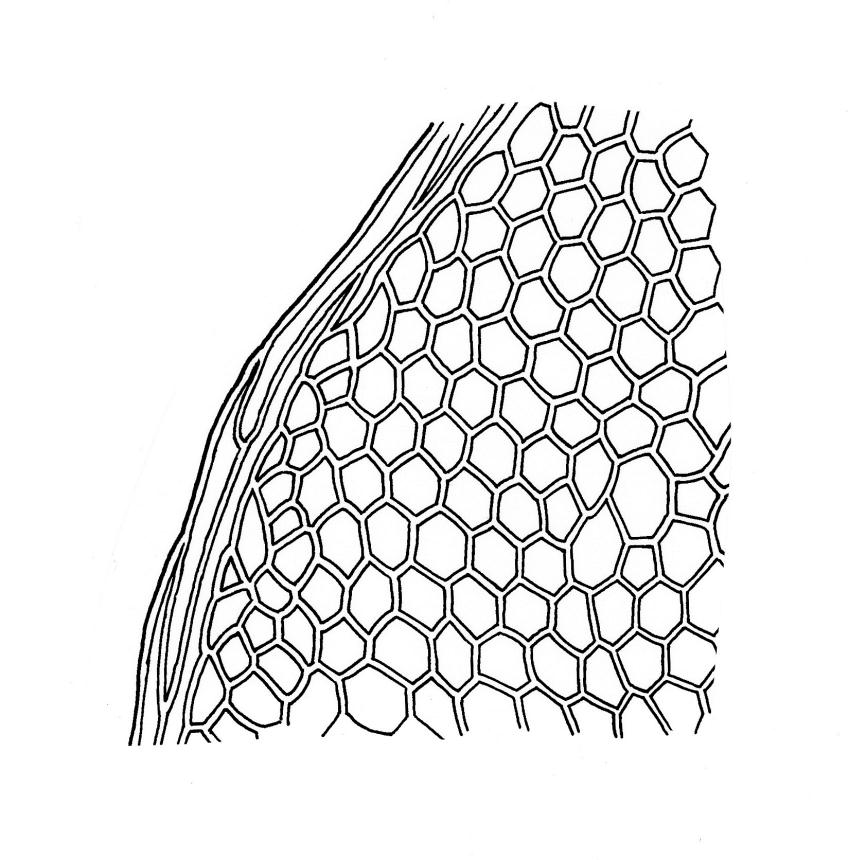
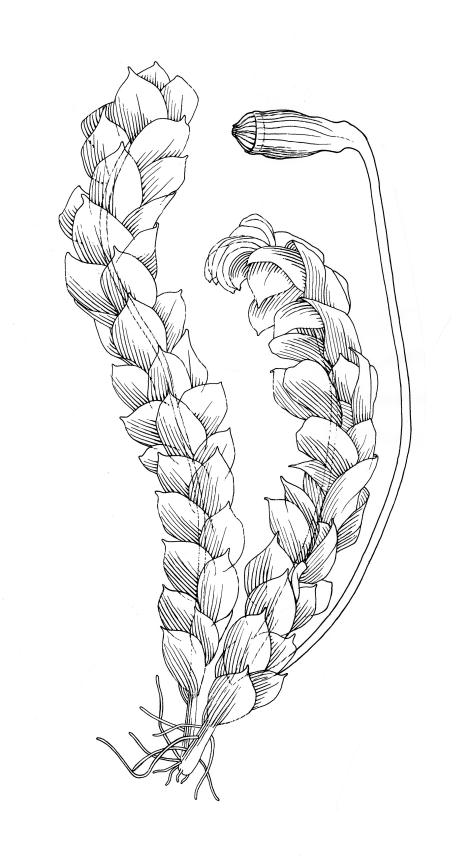
Plants small, yellow-green to bright green, weakly or not iridescent when dry, forming dense, compact and layered mats. Stems moderately and irregularly branched, red-brown, to c. 10 mm (rarely 20 mm). Shoots 1–2 mm wide. Leaves inserted in c. 6 ranks, ± symmetric, weakly complanate, strongly crisped when dry (and then obscuring the complanate nature of the shoot), orbicular-elliptic from a narrowed insertion, strongly apiculate at apex, plane, stoutly bordered, denticulate in upper half or more, occasionally nearly entire; (0.5–)0.7–1.0(–1.3) × 0.3–0.5(–0.6) mm; upper laminal cells firm-walled, rhombic to ± hexagonal, (8–)10–15(–18) × 9–12(–15) µm, becoming larger (to 54 µm in greater dimension), laxer and more oblong towards the base; juxtacostal cells not to moderately differentiated; border of elongate cells stout, extending from insertion to apiculus, c. 15–21 µm wide at mid leaf, concolorous. Costa concolorous, c. 13–18 µm wide below, unbranched, weakly spurred or rarely branched above, extending ⅔–¾ the leaf length and usually ending 200–330 µm below the base of the apiculus.
Dioicous. Perichaetial leaves acute-cuspidate, ecostate, weakly bordered, c. 900 µm long. Perigonia gemmiform, scattered on ♂ stems, lacking paraphyses, with ovate, acute to stoutly acuminate, ecostate, and scarcely bordered bracts c. 500 µm long. Setae 4–10 mm, smooth, weakly sinistrorse, red-brown; capsules inclined to horizontal, oblong-cylindric from a tapered neck, c. 1 mm, constricted below the mouth when dry, pale red-brown; exothecial cells oblong to isodiametric, collenchymatous; annulus not seen; operculum rostrate from a conic base, c. 0.5 mm. Exostome teeth c. 210 µm, otherwise as per genus; endostome segments conspicuous when dry, arising from a high basal membrane, equal to or slightly longer than exostome teeth, perforate. Calyptra mitrate-rostrate, c. 1.0 mm, hairy above. Spores 9–15 µm, green, smooth.
Brotherus 1925, fig. 596; Matteri 1975, pl. 4; Streimann 1999, fig. 13.
Several non-fruiting collections from Taranaki and Nelson L.D. are difficult to confidently assign to either D. rotundifolium or D. crispulum var. crispulum using available features, and suggest that further study is required to clarify the relationship between the two taxa, both of which are dioicous. The endostome segments of the two taxa appear to differ, with those of D. rotundifolium perforate and those of D. crispulum non-perforate. Distichophyllum rotundifolium as conceived here is a smaller plant with shoots ≤2 mm (vs c. 3 mm) wide. The leaves of D. rotundifolium are narrower (0.3–0.5(–0.6) vs 0.7–1.1 mm) and usually shorter (not or scarcely exceeding 1.3 mm vs 1.0–2.3 mm long), but these size ranges are not discrete. Distichophyllum rotundifolium usually has denticulate leaf margins, while D. crispulum leaf margins are invariably entire. The distinction between the two taxa is obscured by some collections (mostly in WELT), but the two taxa are retained here as distinct pending further study. If these taxa were to eventually prove synonymous it is unclear which would have priority (the basionyms of both were published in 1844).
Distichophyllum rotundifolium is sometimes confused with Calyptrochaeta flexicollis; the features that readily separate them are discussed under the latter species.
K (Ravine 8); NI: N Auckland, including offshore islands (TK, LB, GB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough (Haldon Hills, Endeavour Inlet), Canterbury (near Mt Peel Station), Westland, Otago, Southland; St; Ch; A; Sol; C.
Austral. Tasmania*, mainland Australia (Vic.)*. Additionally reported from N.S.W. by Streimann (1999) and from Argentina and Chile by Matteri (1975).
Occurring from lowland forest to the subalpine zone, this species is catholic in its substrate preferences. It commonly grows on logs in a wide range of decay, and on damp soil banks, usually in shaded situations. It also occurs on tree trunks and branches and rarely grows as an epiphyll (sometimes on fern fronds). It can occupy thin soil over a wide variety of rock types (including limestone) and may be found on rocks submerged in swift or cascading (usually small) streams. It is far more common in the wetter parts of the country and occurrences in the drier parts of the South I. are rare. From near sea level to c. 900 m (Mt Taranaki, Taranaki L.D.) on the North I. and from near sea level to c. 1025 m (Hope Range, Nelson L.D.) on the South I.
Distichophyllum rotundifolium is an extremely attractive species occurring on a wide range of substrates. Once its distinctive combination of small size, usually terrestrial habitat, and strongly crisped, orbicular-elliptic, strongly apiculate, and stoutly bordered leaves is known, this species is easily recognised. The marginal toothing is usually barely visible under the hand-lens, and is sometimes absent. The species can develop longer-than-normal stems when growing on rocks in stream beds.
Dixon (1927, pp. 281–282), placed Hookeria amoena Colenso and H. smaragdina Colenso (both published in Trans. & Proc. New Zealand Inst. 18, 1886) in the synonymy of D. rotundifolium. Dixon based his decision concerning H. amoena on his interpretation of Colenso’s description and he did not see type material. He gave no reason for his decision regarding H. smaragdina. No identifiable type material has been available for study (including in BM) but little would be gained by questioning Dixon’s proposed synonymy.