Class
Family
Genus
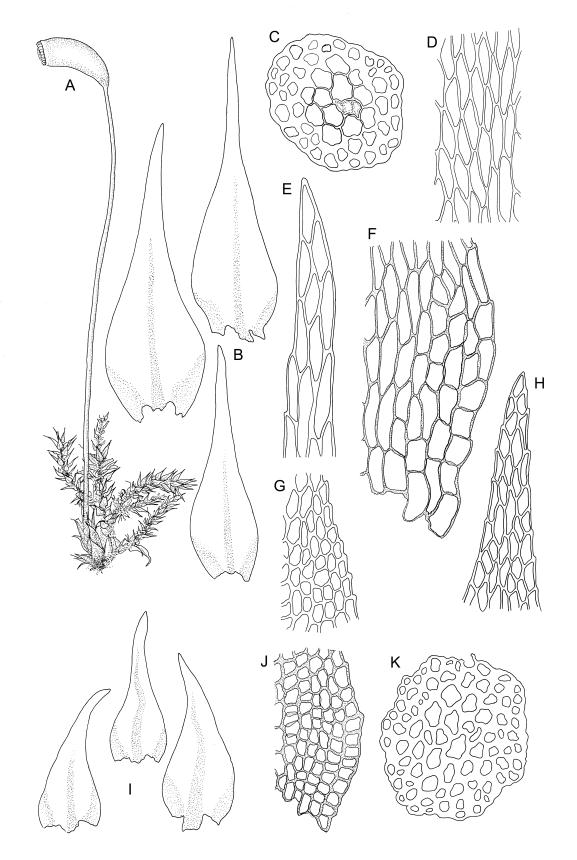
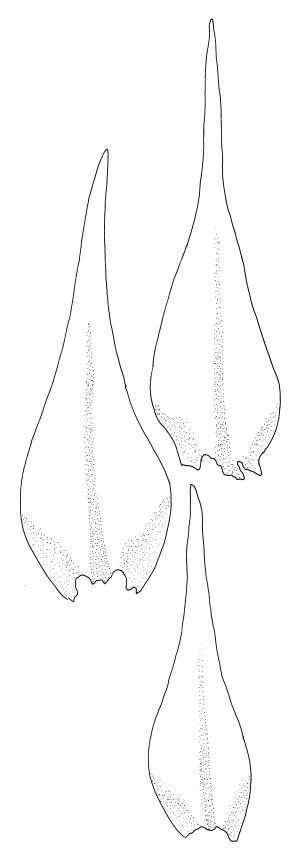
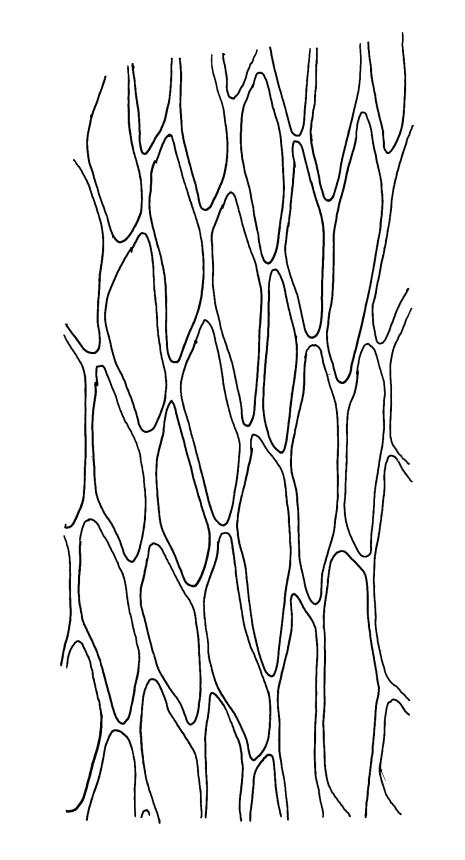
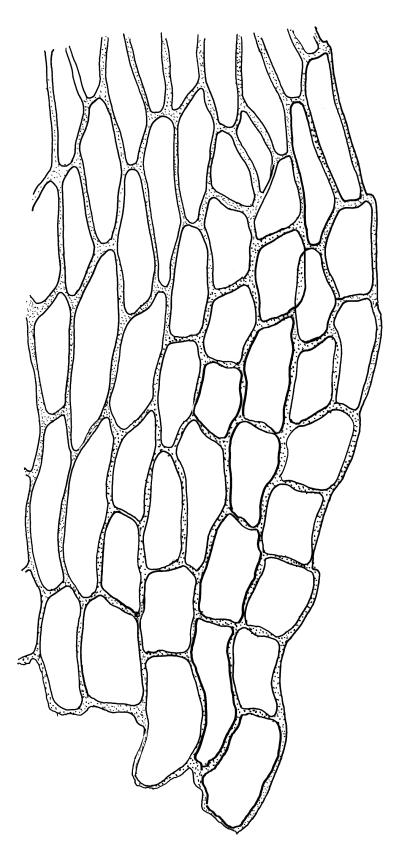
Plants very small, yellow- or bright-green. Stems creeping, in cross-section with c. 3 layers of small, firm-walled cells surrounding larger and thin-walled medullary cells and a small central strand. Stem and branch leaves similar, appressed when dry, spreading to c. 45° when moist, narrowly lanceolate and tapered to a finely acuminate apex, constricted at insertion, weakly denticulate (usually most obviously at mid leaf or below), c. (0.5–)0.6–0.8 × (0.15–)0.2–0.27 mm; mid laminal cells oblong-hexagonal, firm-walled, 24–30 × 7–9 µm and mostly 3–5:1, firm-walled, smooth; alar cells ± quadrate and firm-walled, forming a large but ill-defined group which often extends upwards to beyond the widest part of the leaf. Costa rather ill-defined, c. ½ to ⅔ the leaf length and fading in or near the acumen base. Paraphyllia absent.
Perichaetial leaves lanceolate, c. 1.5 mm. Setae flexuose, rather stout for the size of the plant, c. 12–25 mm; capsule cylindric from a moderately defined neck, strongly curved, c. 1.8–2.2 mm, strongly constricted below the mouth when dry; stomata not seen; operculum bluntly conic. Exostome as per genus; endostomal cilia mostly single or sometimes paired in N.Z. material. Spores 12–15 µm, finely papillose.
Kanda 1976, fig. 6, 1–14; Crum & Anderson 1981, fig. 451; Smith 2004, fig. 252, 1–4.
Amblystegium serpens is most likely confused with those forms of Fabronia australis in which marginal spines are weakly developed. Amblystegium serpens lacks the very fine and "silky" appearance of Fabronia, is not secund, and the alar group is smaller (not extending to the costa base as it frequently does in Fabronia). Both species are autoicous and when fruiting are easily distinguished. Amblystegium is predominantly terrestrial while Fabronia is predominantly epiphytic or epilithic.
Amblystegium serpens is sometimes confused with Rhynchostegium muriculatum but it can be distinguished by its non-complanate habit, shorter mid laminal cells, more strongly differentiated alar groups, smaller leaves, bluntly conic opercula, smooth setae, and other sporophytic differences.
NI: N Auckland (Whangārei, Onehunga, Auckland City), S Auckland (Mayor I.), Hawke’s Bay (Wairoa, Māhia Peninsula), Wellington (Whanganui, Upper Kaumingi Stream, Glenside); SI: Nelson (Mt Arthur, Mt Owen), Canterbury (Woodend, Castle Hill, Broken River), Otago (Pigroot Creek, near Herbert, Mātukituki Valley); Ch.
Bipolar but some N.Z. populations are probably adventive. Widespread in northern hemisphere.
In lowland highly modified sites such as damp soil at drain margins and on rotting wood (e.g., Salix fragilis, Populus alba) in swamps or at the edge of lakes or lagoons. This species also occurs in garden situations, including the soil of flowerpots and in greenhouses. These habitats suggest that the lowland (<c. 100 m a.s.l.) populations may be adventive. However, there are scattered records from less modified and higher elevation habitats where it occurs on soil in moist, and usually calcareous situations. The species is known from c. 300 m at the Mātukituki Valley, c. 600 m from Pigroot Creek, c. 700–750 m at Broken River, and from c. 1650 m on Mt Arthur. At Broken River it is recorded from the base of a limestone boulder with Gymnostomum calcareum, and the Mt Arthur record is from the soil inside a narrow, 3 metre deep alpine doline. These occurrences suggest A. serpens is indigenous, at least in part.
Lowland collections are mixed with a range of sometimes weedy species including Amphidium cyathicarpum, Ceratodon purpureus, Cratoneuron filicinum, Cratoneuropsis relaxa, Eurhynchium praelongum, Leptobryum pyriforme, Plagiomnium novae-zelandiae, and Rhynchostegium muriculatum.
High elevation collections are generally more compact, with leaves more erect-appressed than low elevation populations.