Class
Family
Subfamily
Genus
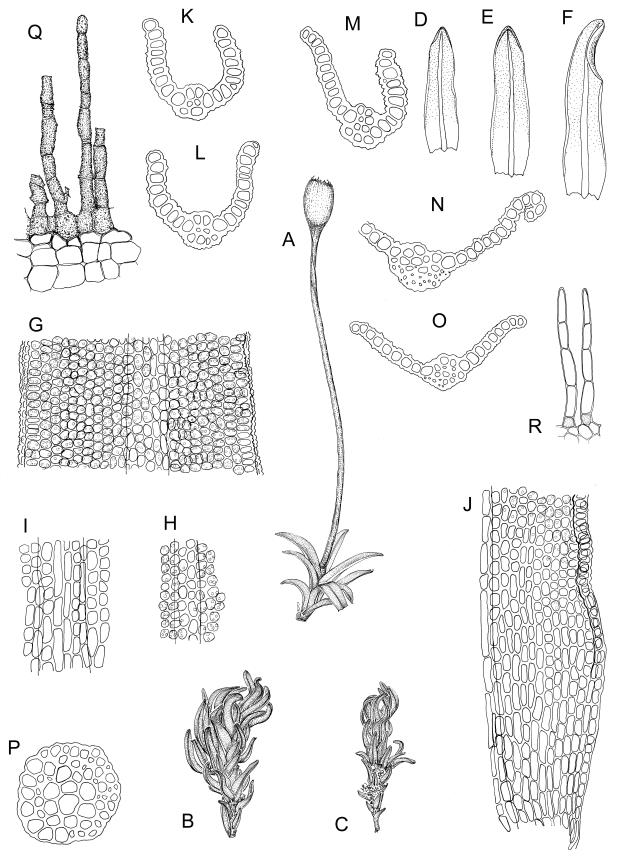
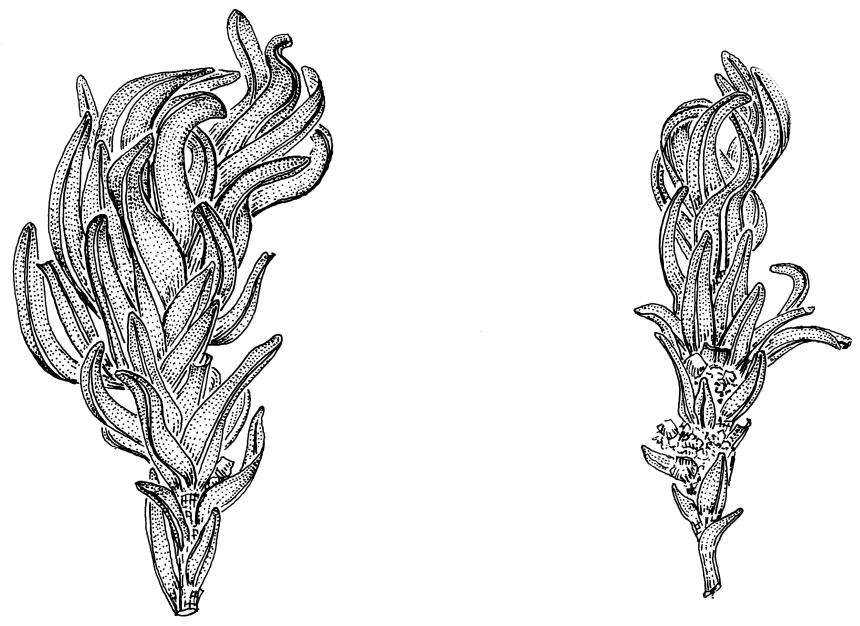
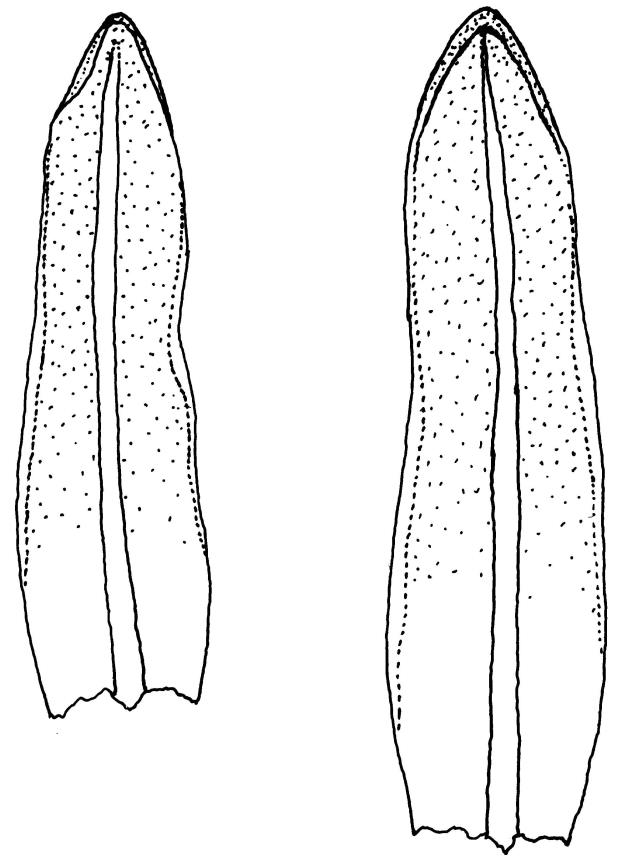
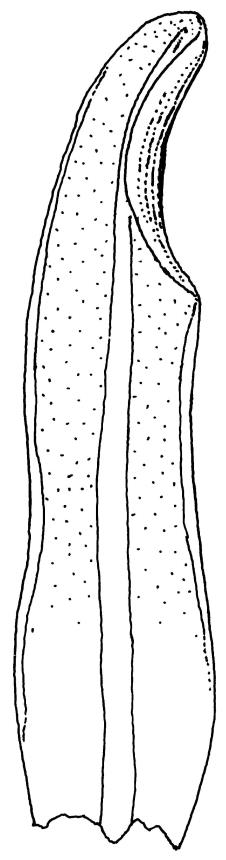
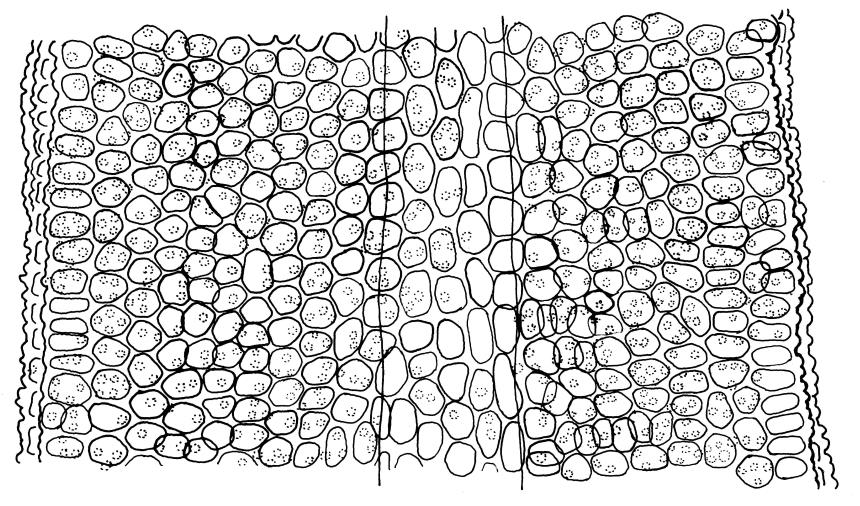
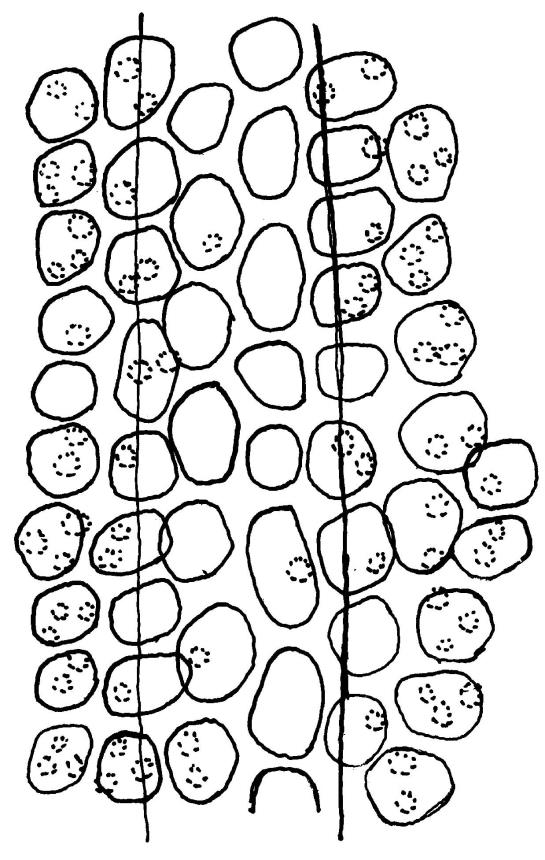
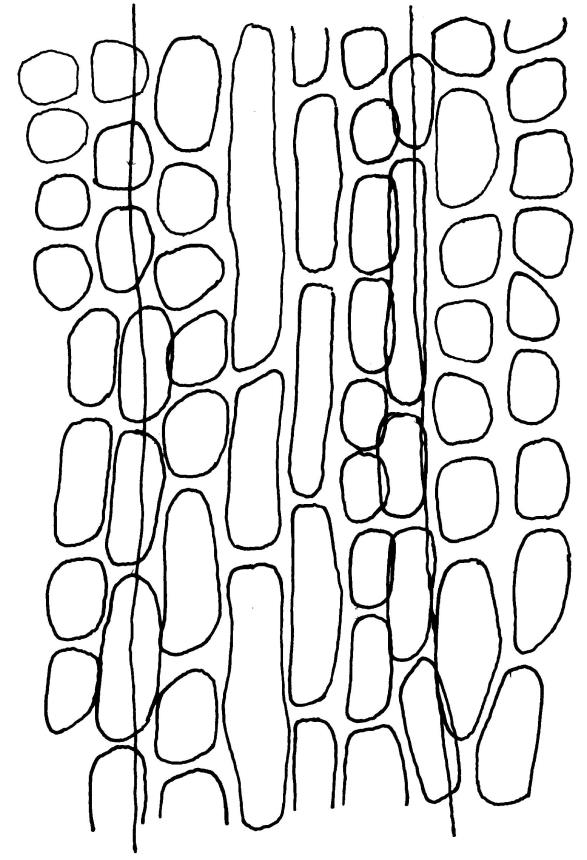
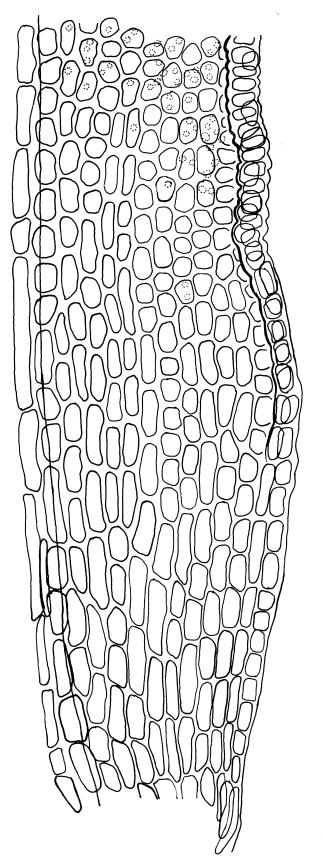
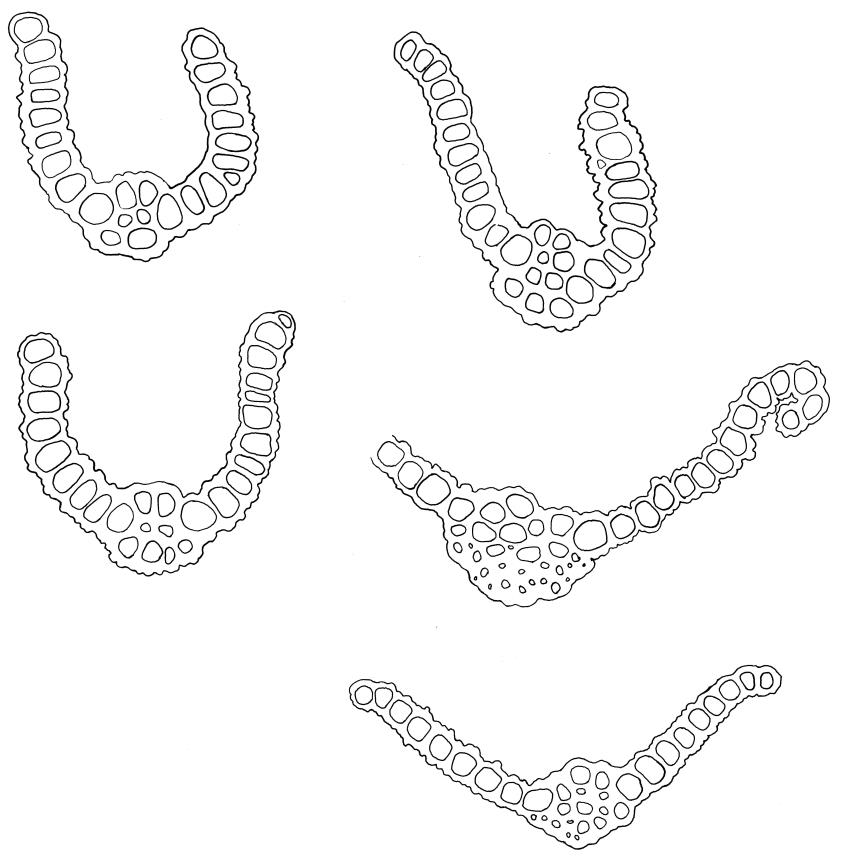
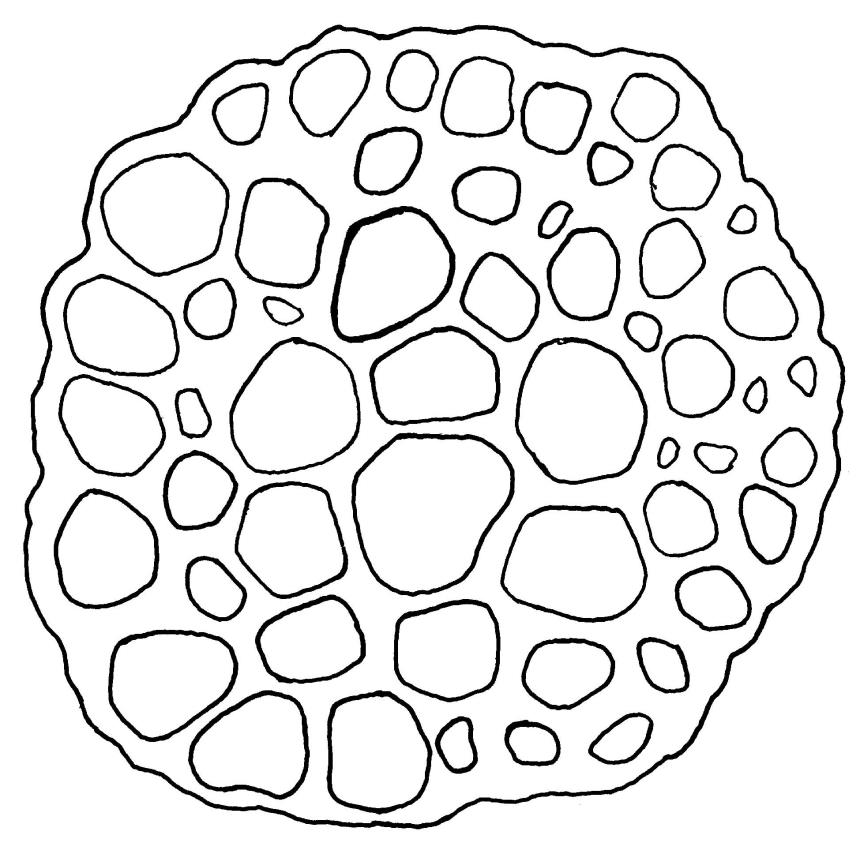
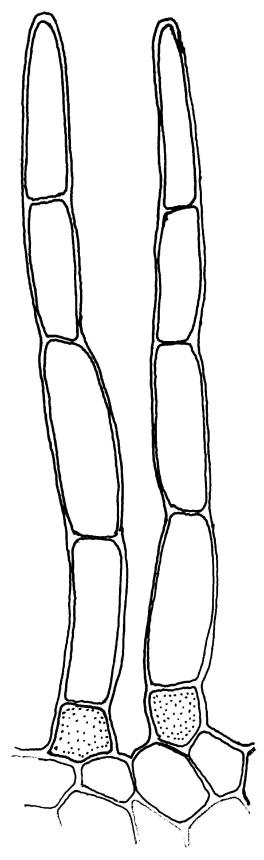
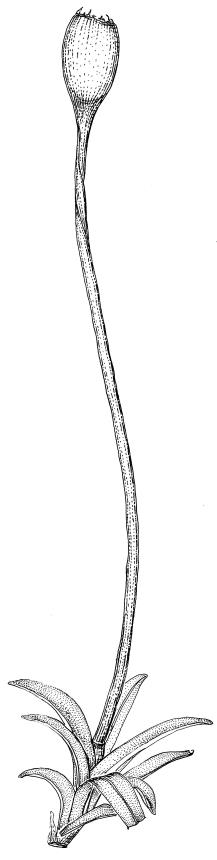
Plants glossy, dark brown-green, bright green in apical leaves only, densely gregarious. Stems 3–10 mm, branched by innovations from below perichaetia and perigonia, in cross-section without a central strand, outer cells thicker-walled and smaller-lumened but not forming a distinct sclerodermis. Leaves mostly recurved when moist, erect-incurved, or stiffly and irregularly curved around the stem but not crisped when dry, c. 0.8–1.5 mm, lingulate, broadly channelled (and often difficult to flatten under a cover-slip), rounded and cucullate (often strongly so) at apex; margins recurved, sometimes only at mid leaf, or on an occasional leaf plane throughout, ± entire but papillose; upper laminal cells clear in outline, ± isodiametric with rounded lumina, often oblate (especially in marginal row), firm- to thick-walled, somewhat bulging, papillose with (1–)2–4(–5) low, rounded, and simple papillae, (7.5–)9–11(–12) × (7.5–)9–11 µm, becoming longer towards leaf base and oblate towards leaf margin, c. 10–15 cells between costa and margin at mid leaf; lower laminal cells weakly differentiated, rectangular, to c. 3 × 1, firm-walled, smooth, shorter at the margin. Costa concolorous, but becoming dark reddish brown in older leaves, stout, failing c. 5–10 cells before the apex; adaxial superficial cells quadrate to short-rectangular in at least distal ⅓, sometimes similar to adjacent laminal cells throughout; abaxial superficial cells quadrate distally. Axillary hairs of 3–4(–6) cells with basal cell shorter and brown. Laminal KOH colour reaction yellow-orange.
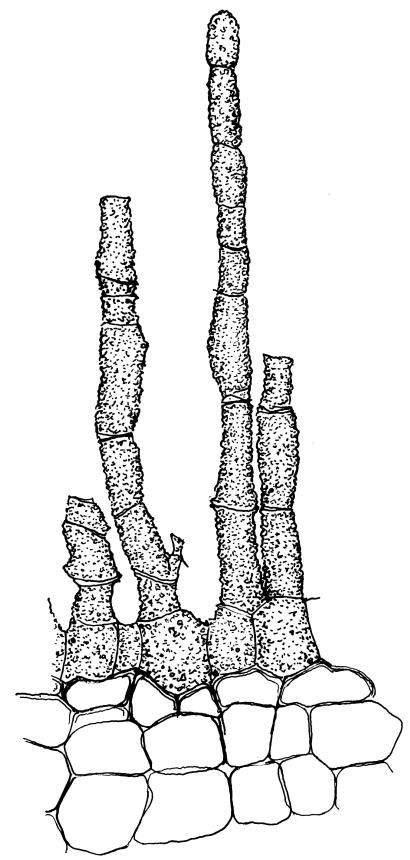
Dioicous. Perichaetia terminal with leaves longer and somewhat broadened at the base, the basal cells elongate, smooth, thin-walled and lax, in a zone extending to c. ⅓ of the leaf length. Perigonia terminal, bulbiform, on an equal-sized plant. Setae red, 4–7 mm. Capsules erect, symmetric, short- or long-ellipsoid, 0.75–1.0 mm. Operculum obliquely rostrate from a conic base, ½ to equal the theca length, with cells in straight rows. Peristome short (c. 200 µm), erect, orange-brown, of 16 densely papillose and articulate teeth, imperfectly formed, occupying only the base of the operculum (with detached distal portions sometimes visible through the operculum wall), irregularly divided, from a low, papillose basal cylinder. Spores (8–)9–12(–18) µm, smooth.
Malcolm et al. 2020, p. 520–523 (incorrectly labelled as "Didymodon novae-zelandiae").
Tridontium milleneri is distinctive in its dark, lingulate leaves that are stiffly incurved and strongly concave when dry, mostly reflexed when moist, with rounded, cucullate apices and a failing costa. Adaxial costal cells are similar to the adjacent laminal cells in at least the distal ⅓ of the leaf. The short, irregular, erect peristome, within a long-beaked operculum, is also characteristic of the species.
Confusion is most likely with related species that have rounded leaf apices and a failing costa, namely Geheebia ceratodontea, G. tophacea, Tridontium cockaynei, and T. novae-zelandiae. All can be distinguished using vegetative characters alone, by means of the key to N.Z. genera in subfamily Barbuloideae, and the key to species of Tridontium.
Mixtures with non-fruiting Gymnostomum calcareum can be puzzling, as shoots of that moss are superficially similar to fresh, bright green growth of T. milleneri. The dark colour of mature shoots of T. milleneri distinguishes them from Gymnostomum calcareum, as do the leaf margins being at least partially recurved (vs plane leaf margins in G. calcareum). As in other species of Tridontium, the rounded leaf apex (vs an occasional leaf apex tapered to a single cell in G. calcareum) is a useful distinguishing character. When plants are fruiting, the pale yellow setae and gymnostomous capsules of G. calcareum will distinguish it (vs red setae, and peristomate capsules in Tridontium milleneri).
From Hyophila novae-seelandiae, another moss of calcareous substrates, T. milleneri differs in the glossy dark green colour of its dry shoots (vs dull reddish brown in H. novae-seelandiae), and its cucullate leaf apices (vs flat or slightly recurved in H. novae-seelandiae). In addition, T. milleneri has c. 10–15 files of laminal cells between costa and margin at mid leaf (vs 20–40 cells in H. novae-seelandiae).
NI: N Auckland, S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson; Ch.
Endemic.
Most collections of this moss have been made in coastal seeps on exposed calcareous substrates, such as limestone or papa rock, and subject to salt spray. It is known also from inland sites, near Taumarunui in Taranaki L.D. at 180 m elevation (P. Beveridge MX-5, 22 Nov. 2013, WELT M041678), and in the Pohangina Valley in Wellington L.D. at 440 m elevation (P. Beveridge EC-10, 13 Dec. 2005, WELT M037168). Although most records are from the North I., the species extends southwards to the west coast of Nelson L.D., and to the Chatham Is. Given the abundance of apparently suitable sites, and the diminutive stature of this moss, many more records are to be expected. Associated mosses include Bryum clavatum, Fissidens leptocladus, Geheebia ceratodontea, and Gymnostomum calcareum.
Confusion has long occurred over the appropriate name for this moss. Specimens have been variously named Desmatodon lingulatus (Hook.f. & Wilson) Sainsbury, Didymodon lingulatus (Hook.f. & Wilson) Broth., Didymodon weymouthii (R.Br.bis) R.H.Zander, and Weissia weymouthii R.Br.bis. Molecular analysis (Beever et al. 2023) has confirmed its position as a new species in the Australasian genus Tridontium. Over 40 specimens of T. milleneri are currently known. Five of these (from three separate gatherings) have been located in Sainsbury’s herbarium at WELT, all previously tentatively identified by Sainsbury as "Weissia weymouthii".