Class
Family
Genus
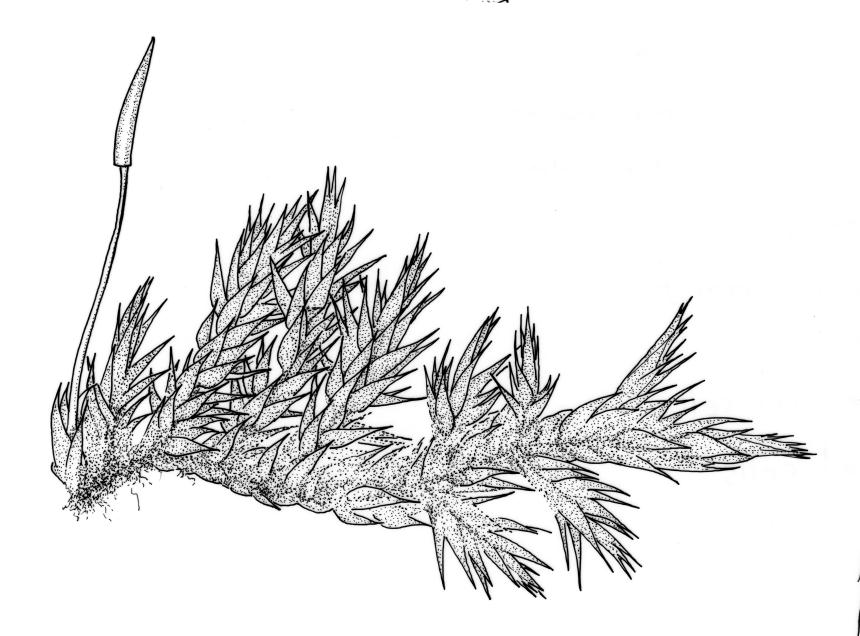
Plants yellow- or white-green, lustrous, ± caespitose, mostly on rotten wood. Stems irregularly branched, <10–15 mm, weakly ascendant, with smooth, brown rhizoids in fascicles on ventral surface, in cross-section lacking a central strand and hyaloderm. Branches short, mostly 3–5 mm. Leaves strongly homomallous, pointing away from, or parallel to, the substrate, narrowly ovate-lanceolate or lanceolate-acuminate, evenly tapered to a slenderly acuminate apex, moderately narrowed to insertion, weakly concave, plane or narrowly reflexed, entire, 0.8–1.3(–1.5) × 0.25–0.4 mm (under cover slip); mid laminal cells (upper third of leaf) linear-rhombic and pointed at ends, mostly 42–75 × 5–7 µm and c. 7–13:1, smooth, not porose; cells at leaf base becoming shorter and porose and a basal band often yellow; alar cells abruptly differentiated, usually 2–4 in extreme angles strongly inflated (c. 45 × 20–30 µm) and hyaline or pale yellow, with supra-alar cells few and irregular or more numerous and extending 4–5 cells up the margin.
Autoicous in N.Z. material. Perichaetia scattered on stems, the inner perichaetial leaves narrowly acuminate from an oblong and ± sheathing base, serrulate above, c. 1.1–1.3 mm. Perigonia small and inconspicuous on stem, with inner bracts broadly ovate and c. 0.35 mm. Setae red at maturity, smooth, twisted weakly to the left when dry, 7–9 mm; capsules horizontal or pendulous due to curvature of setae, oblong-cylindric, with a short and ill-defined neck, ± constricted below the mouth when dry, little altered when moist, pale yellow-brown when mature, 1.0 –1.5 mm; stomata restricted to neck, superficial; annulus not differentiated; operculum slenderly rostrate from a conic base, ± equal to the urn in length; exothecial cells short-oblong or ± quadrate, strongly collenchymatous, not in distinct longitidinal ranks. Peristome as per genus; endostome with a high basal membrane and single nodose cilia. Calyptra cucullate and smooth. Spores 12–18 µm, green, finely papillose.
Confusion can occur between the present taxon and depauperate forms of Wijkia extenuata. Sematophyllum subhumile var. contiguum is generally a paler, more lustrous plant in which the stem leaves are smoothly tapered to the apex, rather than abruptly tapered as is usual in Wijkia. New Zealand material of S. subhumile var. contiguum is autoicous and usually bears at least some fruit, while Wijkia is dioicous and often sterile. In S. subhumile var. contiguum the leaves are entire and the laminal cells invariably smooth while in Wijkia some degree of toothing almost always occurs near the leaf apex and the laminal cells usually exhibit some degree of papillae development. Material of S. subhumile var. contiguum lacks flagelliform branches, while W. extenuata var. caudata usually produces reduced flagelliform branches.
Confusion sometimes occurs between S. subhumile var. contiguum and S. homomallum, but the former generally occurs on wood (not rock), is paler, has more distinctly inflated alar cells and serrulate (not entire) perichaetial leaves. The two taxa also differ by their sexuality in N.Z.
Sematophyllum kirkii is similar in habit to S. subhumile var. contiguum, but differs from it by having subcollenchymatous exothecial cells, mammillose setae, and sharply toothed vegetative leaves.
K; NI: N Auckland including offshore islands (TK, LB, GB, RT, Maria I.), S Auckland, Gisborne (Anaura Bay, Paremata), Hawke’s Bay, Wellington; SI: Nelson (Parapara, Fenian Track, Lake Rotoiti, Punakaikī), Marlborough, Canterbury, Westland (Taramakau), Otago, Southland; St; Ch.
Anomalous. Tasmania*, mainland Australia*. The variety contiguum is recorded from Tasmania, South America and Oceania by Ramsay et al. (2002); they (p. 37) considered S. subhumile s.l. to be "a common species distributed in temperate and tropical Asia, South Africa, the Pacific Islands, New Zealand and Australia also Norfolk Island."
Mostly on rotten wood. Less often on exposed roots or rock (including greywacke) or soil; occasionally epiphytic (on southern beech, Fuchsia excorticata, Podocarpus, Rhopalostylis, and Cordyline. Occurring in a range of native forest types and also in exotic (e.g., Eucalyptus, Populus, and Pinus) plantations. On South I., this species is far more common in eastern regions and is a common and widespread species at lower elevations in Canterbury and Otago. On North I. from near sea level (Bay of Islands, N Auckland L.D.) to c. 860 m (Kaweka Range, Hawke’s Bay L.D.), but with most records at or below c. 400 m. On South I. from near sea level (Christchurch) to 540 m (Island Hill, Canterbury L. D.). Frequently associated bryophyte taxa include Dicranoloma menziesii, Pyrrhobryum mnioides var. contortum, Rhaphidorrhynchium amoenum, and Sauloma tenella.
| Category | Number |
|---|---|
| Indigenous (Non-endemic) | 1 |
| Total | 1 |
For convenience, the varietal name contiguum employed by Ramsay et al. (2002) and long associated with N.Z. and Tasmanian material, is applied here. However, the differences cited by those authors (p. 43) to separate S. subhumile var. subhumile and var. contiguum are not great. They refer to Sainsbury (1955) in making those distinctions but a close reading of Sainsbury indicates that he was sceptical of the separation of these two taxa. According to Ramsay et al. (2002, p. 43), the main difference between these varieties is that var. contiguum has serrulate perichaetial leaves and "more acute" vegetative leaves. Their comments about vegetative leaf apices are difficult to interpret, but their figs 19, a1 and a2 indicate those of the var. contiguum to be narrower and sharply pointed, while those of the var. subhumile are broader and merely acute. However, in N.Z. material there appears to be continuous variation in vegetative leaf apical form. Further, I consider the holotype of the Australian Sematophyllum aciculum Broth. ex Dixon to be indistinguishable from N.Z. material. Ramsay et al. (2002, p. 37) interpreted S. aciculum as a taxonomic synonym of S. subhumile var. subhumile. The distinction between the two varieties is blurred at best.
Several N.Z. collections are clearly autoicous. Unless there is a difference of sexuality between Australian and N.Z. populations, the description by Ramsay et al. (2002, p. 37) of the sexuality of S. subhumile s.l. is likely incorrect; dioicy is a rare condition in Sematophyllum.