Class
Family
Genus
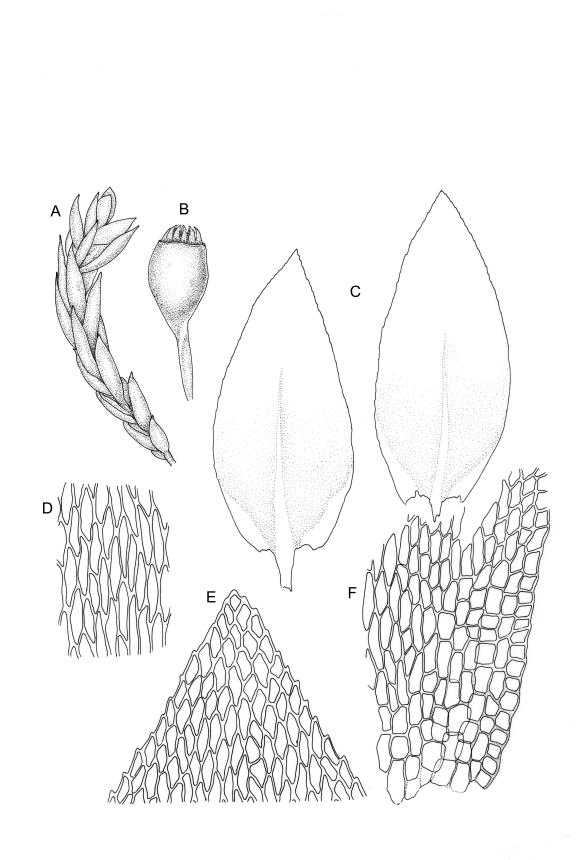
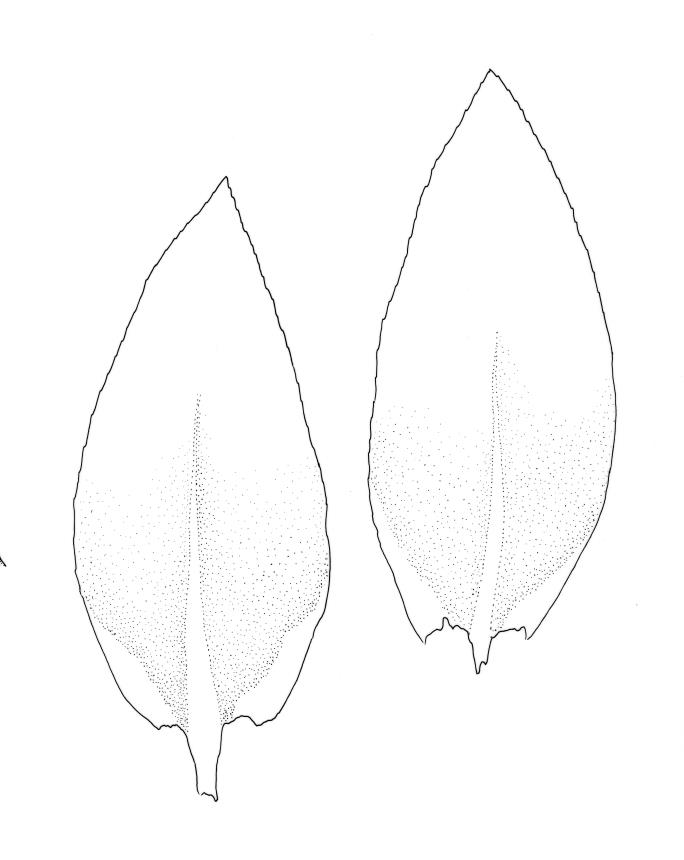
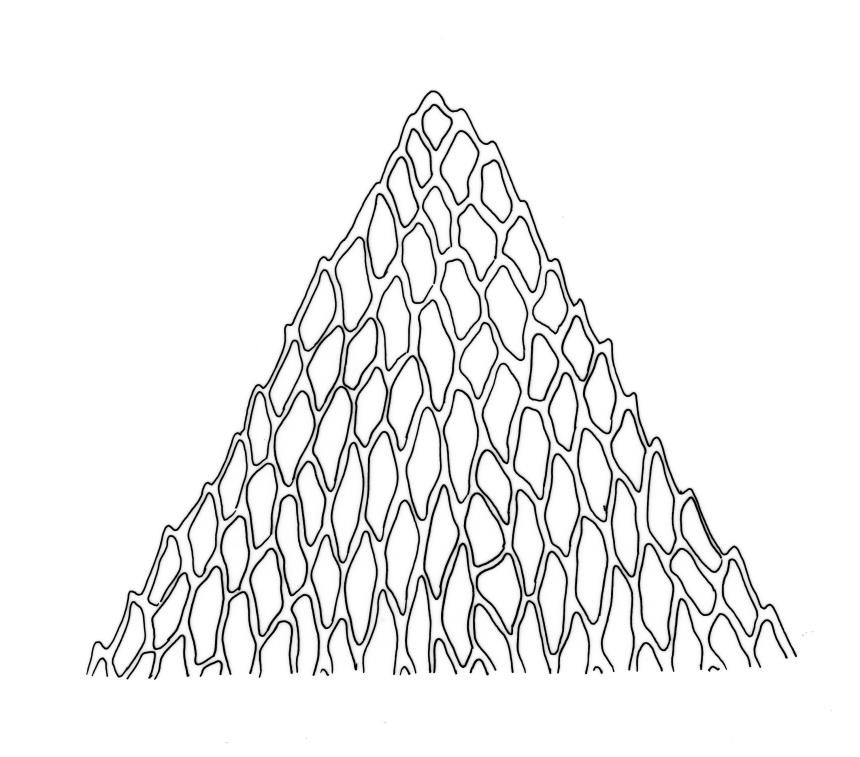
Plants small, very pale or dirty green, dull, prostrate or with branches weakly ascendant and curved, forming densely interwoven mats. Primary stems creeping. Secondary stems mostly c. 10–15(–20) mm long, fastigiately or subpinnately branched, in cross-section oval, with several (c. 5) layers of thick-walled outer cells and a distinct central strand; rhizoids smooth, brown, very sparse, scattered in fascicles on ventral surface of stems. Stem and branch leaves differentiated. Stem leaves not clearly seen in N.Z. material. Branch leaves erect and subtubulose when dry, loosely spreading and concave when moist, elliptic-ovate, widest at or near mid leaf, broadly acute, obtuse, or rounded (usually variable on one plant), mostly 0.6–0.8(–0.9) × c. 0.35–0.4 mm, neither decurrent nor plicate, bluntly denticulate in upper one-fourth or slightly more; upper laminal cells (beyond costa) and cells near margins rhombic, firm-walled and smooth, 21–30 × c. 9 µm; mid laminal cells more elongate, (c. 35–45 µm); alar cells quadrate or subquadrate, firm-walled, forming a very large but ill-defined group extending nearly to the costa and upwards on the margin for c. 150–225 µm and c. 15–20 cells. Costa c. 30 µm wide near base, tapered, sometimes weakly spurred above, extending c. ⅔ to apex, apparently without a terminal spine.
Sexuality as per genus. Perigonia scattered, c. 0.6 mm. Setae stout, 4–5 mm, yellow-brown, weakly flexuose, weakly papillose throughout (visible only under compound microscope), or smooth above; capsule obovoid, weakly inclined, yellow-brown at maturity, not narrowed at mouth, c. 1.1 × 0.65 mm; exothecial cells oblong, firm-walled, not thickened at corners; stomata few at capsule base; annulus and operculum not seen. Exostome teeth yellow-brown, broadly lanceolate, mostly blunt at apex, apparently bordered, transversely striate below, trabeculate on inner surface, c. 210–240 µm; endostome present; segments nearly the height of the teeth and narrowly perforate, arising from a high membrane; cilia apparently absent. Calyptra not seen. Spores mostly 20–24 µm, not smooth (surface details not seen).
Scott & Stone 1976, pl. 79; Hedenäs 2002, fig. 2 a–c.
The diagnosis of Hedenäs (2002) in his Australian Brachytheciaceae treatment applies equally well to N.Z. material: “Scorpiurium cucullatum is easily separated from other Australian species of the Brachytheciaceae by its small size, its large alar groups that extend far up along the leaf margins and consisting of numerous shortly oblate to rectangular cells, and by the strongly inrolled leaf margins when dry.” The very pale coloration is helpful in the recognition of this species.
Because of the curvature of the dry branches and its epiphytic habit, this species could possibly be confused with Leptodon smithii, and in particular with those forms producing microphyllous branches. The secondary stem and branch leaves of S. cucullatum are narrower in relation to their width than the branch leaves of well-developed L. smithii. However, with microphyllous forms of Leptodon, difficulties may be greater. The mid laminal cells are longer and more distinctly rhombic in Scorpiurium than in Leptodon (c. 35–45 µm vs <15 µm). Paraphyllia are lacking in Scorpiurium, while the distinctive strap-shaped paraphyllia of Leptodon smithii are present even in microphyllous forms. The host tree species, pale coloration, autoicous sexuality, and numerous sporophyte characters of Scorpiurium will also help differentiate it from Leptodon.
NI: N Auckland (Kawakawa), Wellington (Upper Kaumingi Stream); SI: Marlborough (Rai Valley).
Australasian. Tasmania*, mainland Australia (Victoria)*. Recorded from Queensland and N.S.W. by both Scott & Stone (1976) and Hedenäs (2002).
One of the most poorly documented species in the N.Z. moss flora. The Kawakawa collection was recorded by Bartlett (1984, as Eurhynchium), and this appears to be the only N.Z. published record. This collection is ample and sparsely fruiting; it came from “base of willows on silt (from flooding)”. Bartlett (1984) recorded some nine associated moss taxa, of which Cyptodon dilatatus was the most abundant. Both the Rai Valley collection (from “Browns Road Swamp” and collected by J.K. Bartlett) and the Upper Kaumingi Stream collection (on Te Kanuka Farm Station, collected by P.J. de Lange) also came from silt-covered willow bases. The former was associated with Hennediella serrulata and the latter with Amblystegium serpens. The habitat for the documented collections is very similar to the “typical habitat” of the species in Australia, as described by Scott & Stone (1976, p. 428).
The original Rai Valley collection was made in 1979 but an attempt to relocate this species there in October 2004 was unsuccessful; the silt-covered tree (willow) trunks occupied by this species are possibly an ephemeral habitat at any one site.