Class
Family
Genus
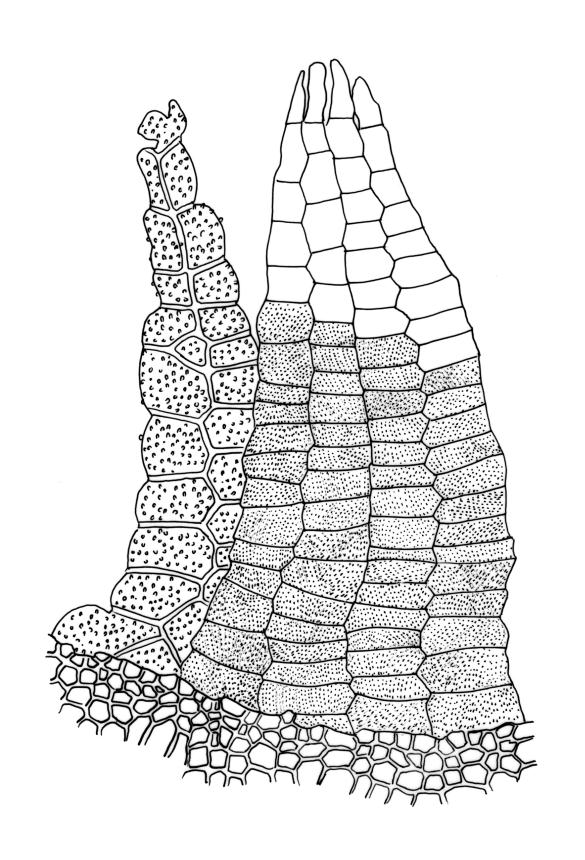
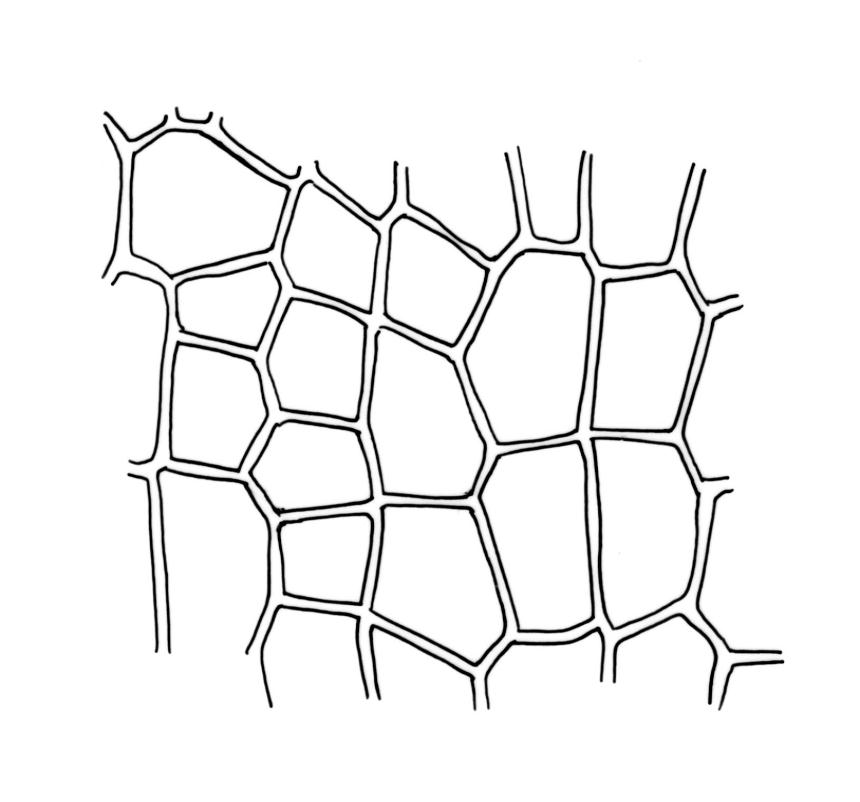
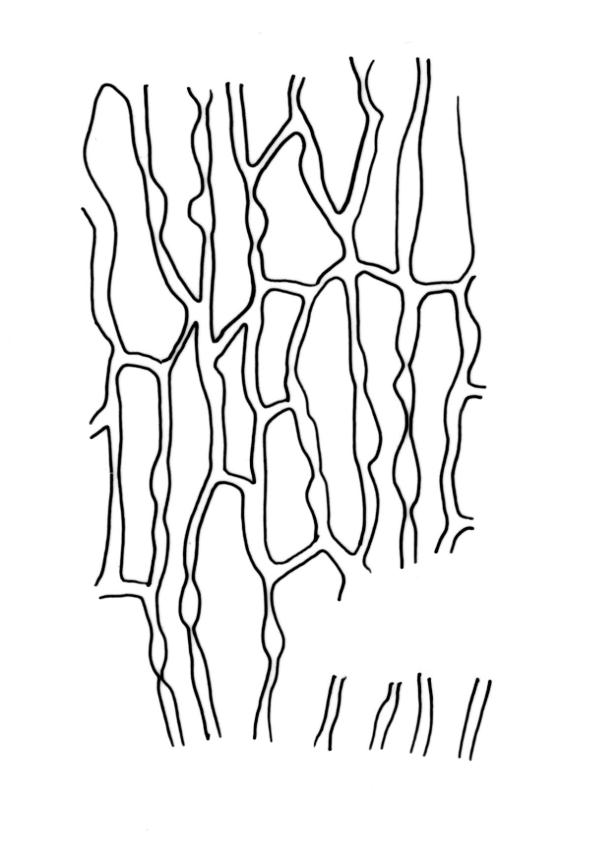
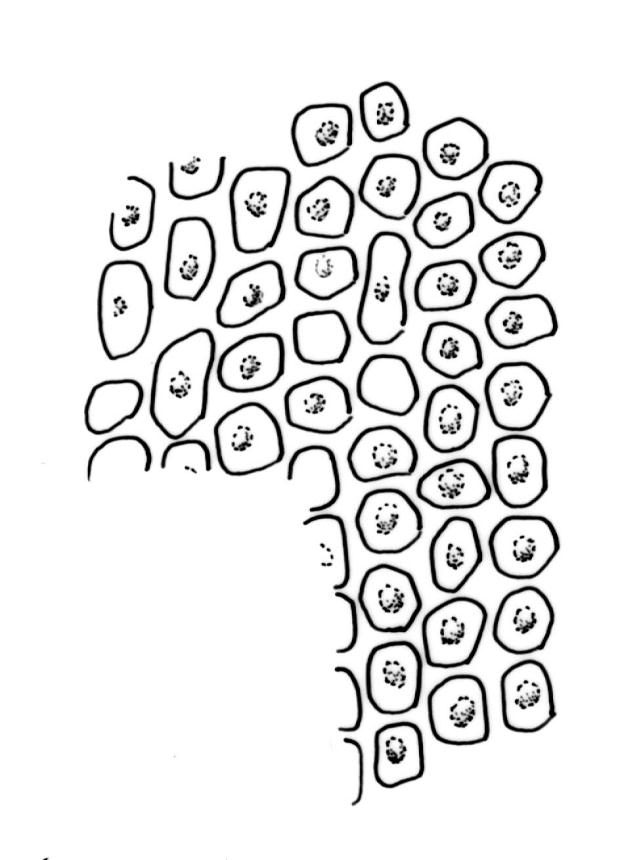
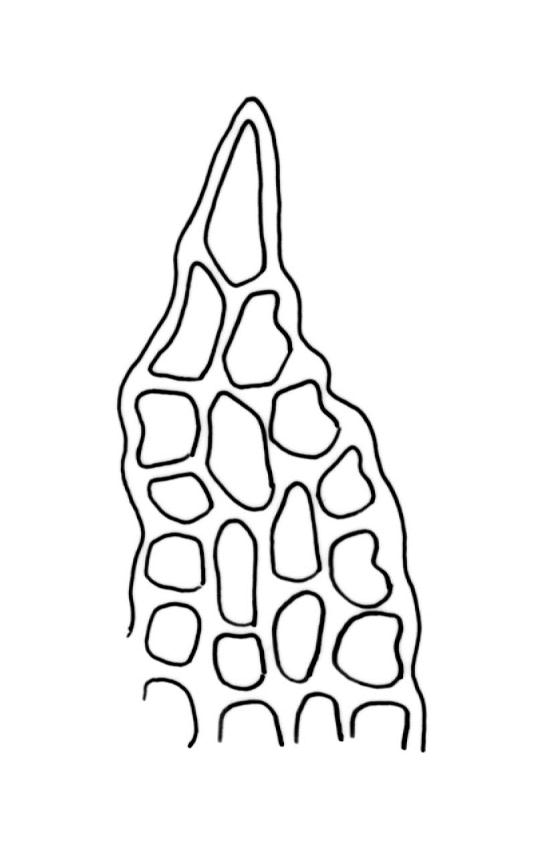
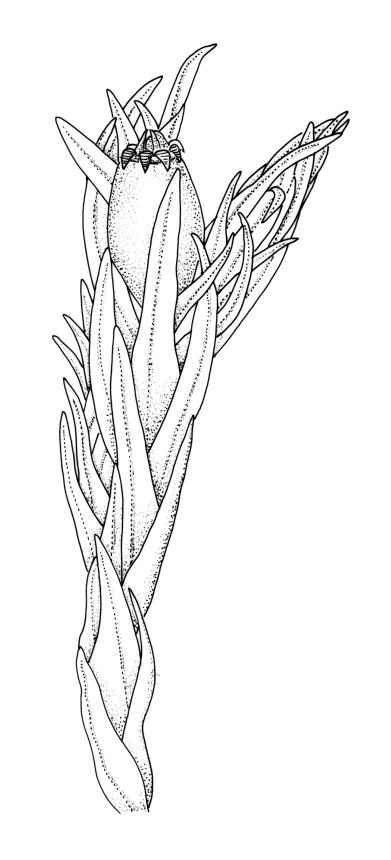
Plants (10–)25–40 mm, forming loose tufts or mats, brown-yellow to olive-green above, darker below. Stems creeping or suberect, much-branched by innovation, flexuose, beset on older parts with abundant smooth, brown rhizoids. Leaves erect-appressed and straight or slightly twisted when dry, erect-spreading when moist, ovate-lanceolate, weakly plicate and sometimes slightly auriculate near the base, acuminate, entire, plane or recurved in the central part or below on one or both sides, 2.6–3.7(–4.1) × 0.7–1.1(–1.3) mm; mid and upper laminal cells isodiametric, oblong, or ± irregular, usually rounded, incrassate, unistratose, (6.0–)9–19(–24) × 6.5–13 µm, papillose with 1–2(–3) low, unbranched papillae per cell; basal interior cells elongate or ± rectangular, thick-walled, nodose, and porose, mostly orange-brown, (30–)51–60(–90) × 6–12 µm, with few to several rows at margins shorter, more regularly rectangular and with thickened transverse walls. Costa as per genus. Gemmae absent.
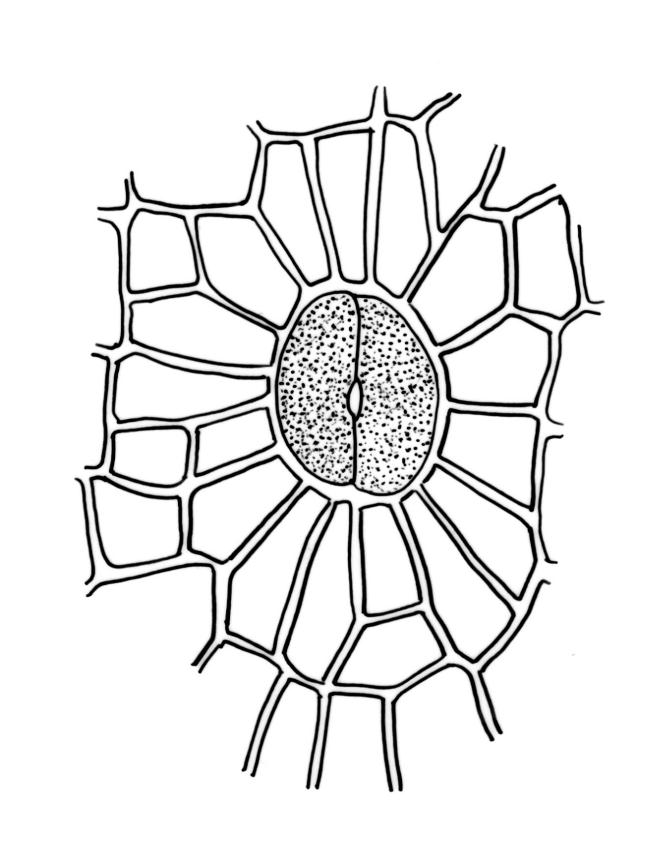
Gonioautoicous. Perichaetial leaves ± enlarged, c. 4 mm. Perigonia gemmiform, conspicuous below the perichaetia and on lower stem. Setae <1 mm; capsules immersed, broadly ovoid or broadly ellipsoid, pale yellow, smooth and constricted at mouth when dry, smooth and not furrowed when moist, 1 or occasionally 2 per perichaetium, persistent, with the older appearing laterally on stem; exothecial cells thin-walled, not differentiated into bands, mostly rounded-oblong or oblong-hexagonal; stomata superficial, numerous in the mid and upper part of the urn, often surrounded by a ring of radiating cells; operculum as per genus. Peristome double; exostome teeth 8, pale yellow or orange, strongly or weakly recurved (occasionally ± erect) when dry, the outer surface with dense, branched papillae, the inner surface with longitudinal papillose ridges; preperistome absent; endostome well-developed, when intact as tall as the exostome and forming a cone when dry, often broken in mature material, with 8 hyaline segments, each of 2 rows of cells, sometimes narrowed near base. Calyptra relatively small (covering c. ½ the capsule), golden-yellow, plicate and split at the base, densely covered with yellow hairs that extend past the calyptra apex. Spores globose, (18–)20–26(–32) µm, finely papillose.
Lewinsky 1984, fig. 19. The illustration in Sainsbury (1955, pl. 34, fig. 2) is misleading.
Orthotrichum graphiomitrium is easily recognised by its short-ovoid and completely smooth capsules, which are immersed in the perichaetial leaves. The stems are commonly in excess of 25 mm and the plants form extensive mats of up to 150 mm or greater diameter; the robustness of the plants facilitates the distinction of this from other N.Z. species of Orthotrichum. Apart from variation in stature, the gametophytes exhibit little variability.
Orthotrichum graphiomitrium is often similar in size and by its semi-creeping habit to O. rupestre. Both species often have numerous perichaetia on much-innovated shoots, with each perichaetium bearing a single, weakly emergent capsule. In O. graphiomitrium, the dry mature capsules lack furrows, the exostome is variably reflexed, and its teeth lack a preperistome. The endostome is well developed and observable with a hand-lens in dry material (unless fractured) and the superficial stomata are numerous and distributed in the upper half of the urn. The plants are nearly always epiphytic. By contrast, in O. rupestre, the dry mature capsules have furrows and the exostome is erect-spreading when dry. The exostome teeth have a well-developed preperistome. The endostome can be present or absent, and the superficial stomata sparse and mostly confined to the lower half of the capsule. The plants are largely confined to rock.
Confusion with species of Macromitrium (because of the weakly creeping stems) seems less likely. Branching of the stems is here innovative (associated with perichaetia), while in Macromitrium it is not. The frequently produced sessile capsules, the erect-appressed dry branch leaves, the well-developed endostome, and the distribution of stomata over the urn in O. graphiomitrium should likewise prevent confusion with any Macromitrium.
NI: S Auckland ("Moerangi–Mangakino turn-off"), Wellington (Taihape); SI: Nelson (Lake Peel), Canterbury (Ben More, Mt Torlesse, Arthur’s Pass, near Mt Cook Village), Westland (near Mt Davy, Ōtira Gorge, Franz Josef), Otago (Dunedin area), Southland (Rob Roy Glacier, near Mossburn, Te Ānau, near Wilmot Pass, Homer Tunnel).
Endemic.
Epiphytic on trees and shrubs in areas of high rainfall. Best developed and most common on Hoheria glabrata in wet, subalpine areas, but also recorded by Lewinsky (1984, table 1) from Aristotelia, Carmichaelia, Discaria, and Nothofagus and the introduced genera Populus, Quercus, and Salix. Very rarely occurring on rocks. Usually growing with Calyptopogon mnioides, and less often with Orthotrichum tasmanicum and Sauloma tenella; nearly pure collections of this species are common. Occurring mainly in areas with high rainfall (>150 cm per year), but rarely extending into drier areas. Occurring from c. 200 (near Lake Te Ānau) to 920 m (at Arthur’s Pass).