Class
Family
Genus
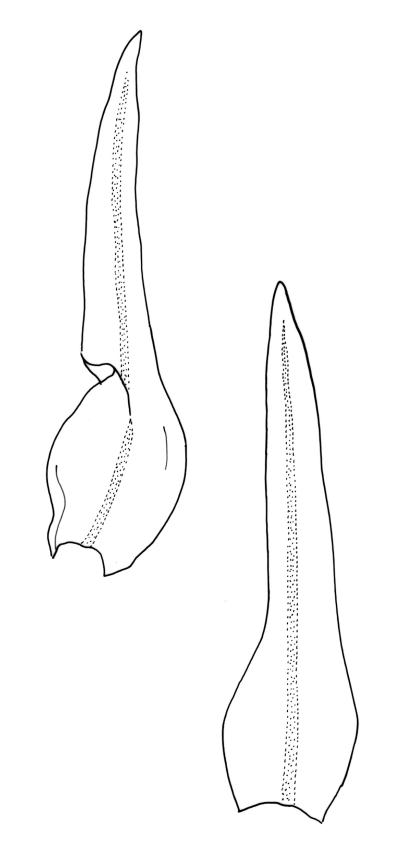
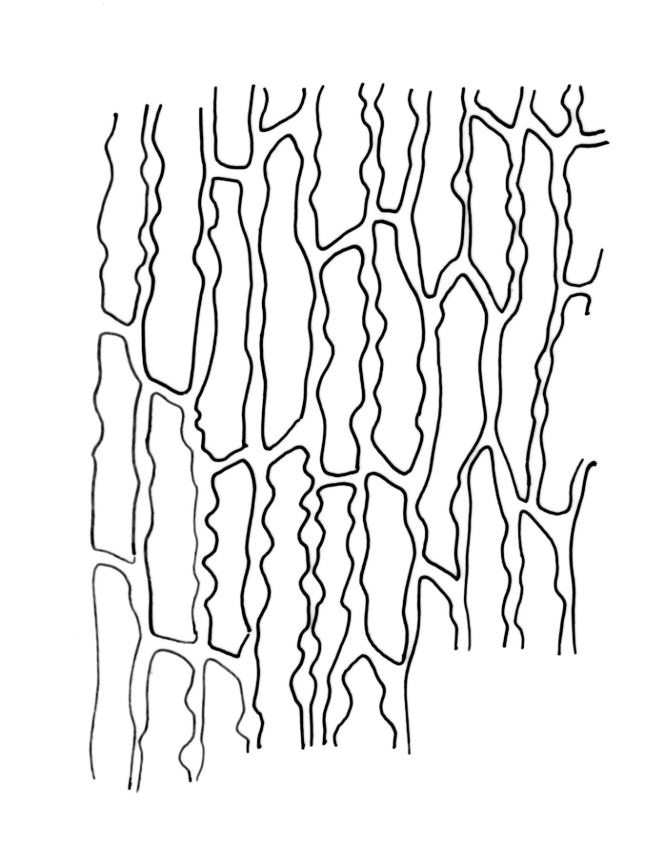
Plants c. 10–30 mm, forming loose or dense tufts, olive-green to bright green or yellow-green above, brown to almost black below. Stems much branched by innovation and by dichotomy, with abundant smooth and brown rhizoids on older parts. Leaves spreading when moist, ± moderately crisped and contorted when dry, lanceolate to ovate-lanceolate, narrowly acute, entire, recurved on both sides from near base to near apex, 2.4–3.5 × c. 0.7–0.8 mm; mid and upper laminal cells mostly angular-isodiametric to ± oblong, occasionally short- to long-rectangular, incrassate, unistratose, 8–14 × (6.5–)8–13 µm, with 1–2 low, branched or unbranched papillae per cell; basal laminal cells elongate, mostly rectangular, some ± rhomboidal, sometimes tapered at ends, thin-walled or incrassate, usually markedly nodose and porose, yellow or yellow-brown, 24–66(–75) µm, with 3–4 rows of cells at margins short-rectangular and with ± thickened transverse walls, the outermost row sometimes continuing far up the leaf margin. Costa as per genus. Gemmae absent.
Gonioautoicous. Perichaetial leaves not differentiated. Perigonia as per genus. Setae c. 3–4(–8) mm; capsules exserted (or emergent or rarely immersed in var. parvithecum), cylindric-ovoid, pale yellow-brown, strongly ribbed in the upper half or smooth (± smooth in var. parvithecum) and slightly constricted below mouth when dry, smooth or nearly so when moist, 1–5 per perichaetium, often of various ages persisting on a single shoot, c. 1.5–2.2 mm; exothecial cells elongate, weakly differentiated into 8 ± pigmented bands, each 3–4 cells wide and extending c. ½ the length of the urn; stomata superficial, restricted to the middle or upper half of urn, sometimes surrounded by a ring of radiating cells; operculum as per genus. Peristome double; exostome teeth 8 (each composed of two paired teeth, and each of the 8 teeth appearing composed of 4 columns of plates), pale yellow to deep orange, c. 300 µm × 150 µm, strongly recurved when dry, the outer surface densely covered by small, transversely oriented papillae that sometimes fuse near the base into transverse wavy lines, the inner surface with dense, longitudinally orientated papillose ridges or striae; preperistome absent; endostome with 8 hyaline segments ± equal to the teeth in length, each c. 60–75 µm wide near base and composed of two rows of cells, with a straight or zig-zag median line, appearing papillose. Calyptra campanulate-mitrate, covering c. ⅔ of the capsule, pleated, pale with a red-brown apex, hairy or seldom naked, the hairs smooth or somewhat papillose by protruding cell ends, yellow or hyaline, reaching ± to the top of calyptra. Spores globose, isosporous and 1-celled, 18–24(–32) µm, papillose.
Orthotrichum tasmanicum is probably the most variable of the Australasian Orthotrichum species. Sainsbury (1955) discussed at length the difficulties he had in trying to distinguish it from other described species, and placed O. lancifolium and O. beckettii in synonymy. Lewinsky (1984) re-examined all the relevant material, concurred with Sainsbury’s conclusions regarding those species and, partly following Dixon (1926, p. 176), added five more validly published N.Z. names and several illegitimate ones (including Orthotrichum nudum R.Br.bis., Orthotrichum subulatum R.Br.bis., Orthotrichum curvatum R.Br.bis., all hom. illeg.) to the synonym list. She also placed several Tasmanian and mainland Australian names here. Many of the names regarded as synonyms by Sainsbury and Lewinsky were based on differences of calyptra hairiness, degree of capsule ribbing, and exostome colour.
| Category | Number |
|---|---|
| Indigenous (Endemic) | 1 |
| Indigenous (Non-endemic) | 1 |
| Total | 2 |
After examining several correlated features of sporophyte morphology, Lewinsky (1984, pp. 387–393) reached the taxonomic decision that material with multiple capsules per perichaetium, shorter setae, and generally smaller and less distinctly ribbed capsules deserved segregation as the var. parvithecum. Her decision is followed here, albeit with reservations. The difficulty in defining the varieties is discussed in greater detail below.
Distribution and ecology as for varieties.