Class
Family
Genus
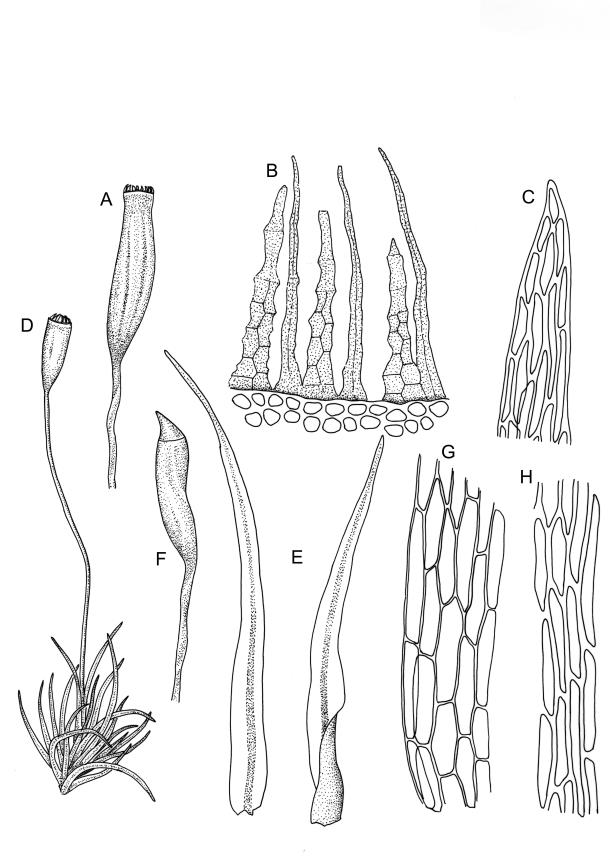
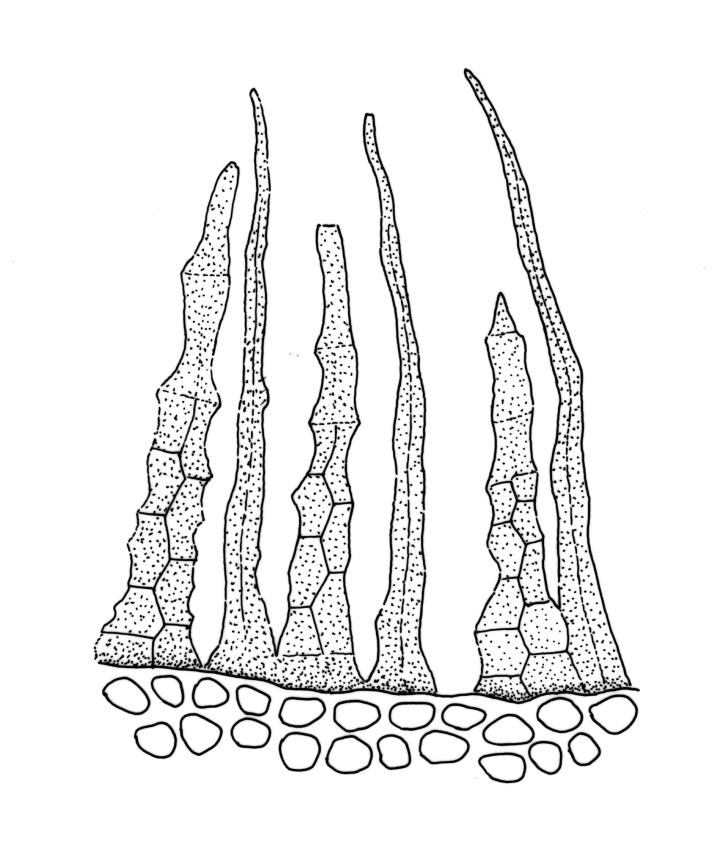
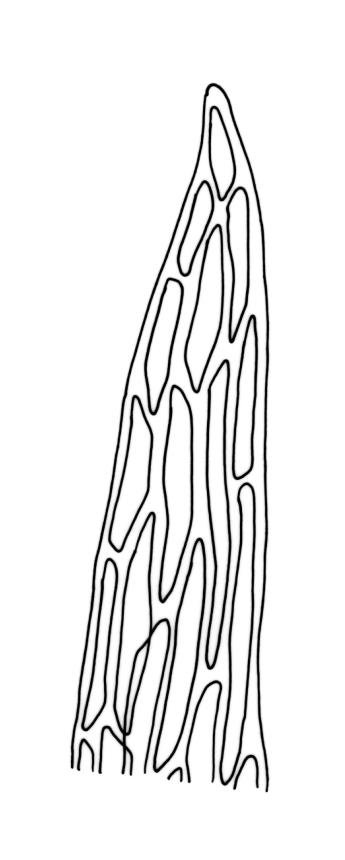
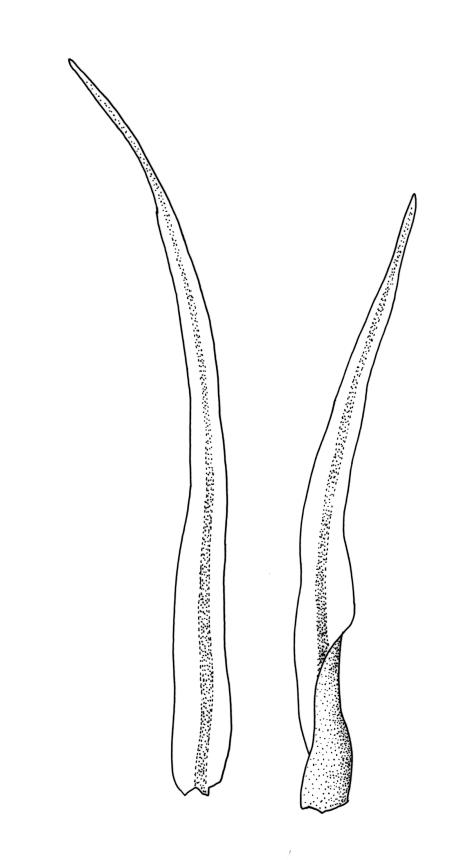
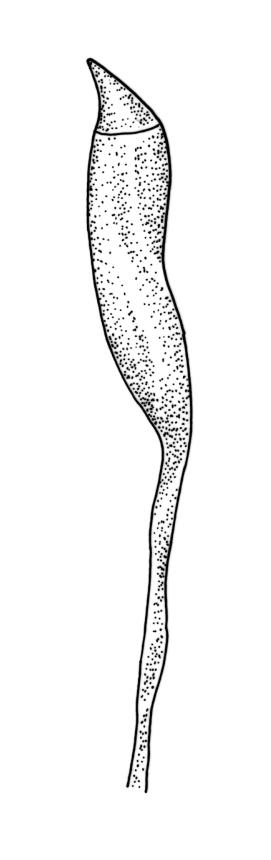
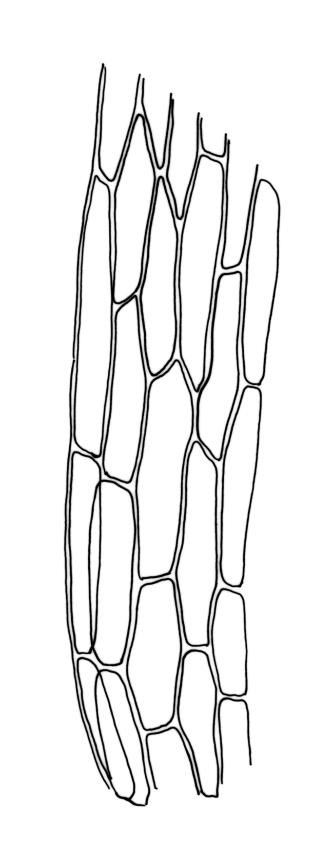
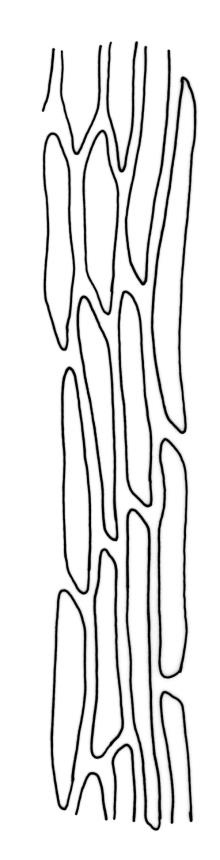
Plants bright- or yellow-green, or gold-brown, forming turves or cushions. Stems very short, mostly <2 mm, unbranched or branching by subperichaetial innovation, beset below with smooth, brown rhizoids, in cross-section with central strand weak or absent. Leaves strongly crowded at stem apex, widely and loosely spreading moist or dry, sometimes weakly secund, flexuose, linear-lanceolate, c. 3–4.5(–6) mm, concave, entire and plane at margins. Mid laminal cells linear, firm-walled, smooth, mostly 90–180 × c. 6 µm, becoming shorter towards apex, and wider, more oblong, and thinner-walled at leaf base; marginal cells not differentiated; alar cells scarcely differentiated. Costa percurrent or subpercurrent, filling <⅕ of the leaf base, in cross-section with a central stereid band and larger cells exposed on both surfaces. Brood bodies (reduced and falcate leaves) occasional on sterile plants; tubers absent.
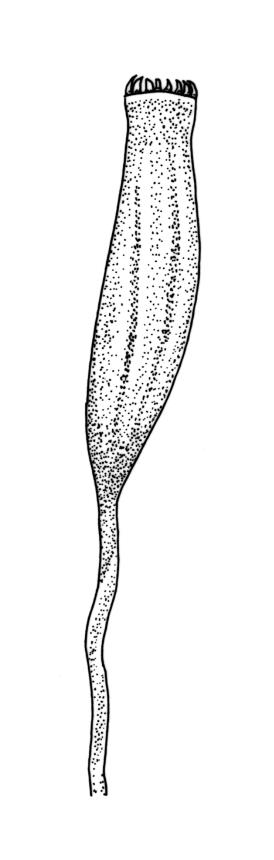
Polygamous (mostly paroicous or gonioautoicous). Perichaetia terminal, with leaves little differentiated. Antheridia usually in axils of outer perichaetial leaves (paroicous) or on short buds below the perichaetium, at the base of fruiting shoots (gonioautoicous), or sometimes terminal. Setae (4–)6–15(–20) mm, pale brown, rather slender, flexuose; capsules erect or inclined, obovoid-cylindric, pale brown or straw-coloured (1.5–)2.0–2.5(–3.0) × c. 0.75 mm, with a short ill-defined neck, weakly to clearly sulcate (8-ridged) and weakly constricted below the transverse mouth when dry; stomata superficial; annulus absent; operculum obliquely rostrate from a conic base, c. 0.5 mm long. Peristome double, fragile; exostome teeth linear-lanceolate, often rounded apically, yellow-brown, strongly inrolled and inconspicuous when dry, erect when moist, trabeculate, slightly papillose, with a nearly straight divisural line; endostome segments pale, longer than teeth (commonly 180–250 µm long), conspicuous when dry, linear, with bulging nodes, finely papillose, with no basal membrane or cilia. Calyptra small and cucullate. Spores spherical, (16–)18–22 µm, papillose-insulate.
Meijer 1952, pl. VIII (as O. lineare var. sulcatum); Seppelt 2004, fig. 80
The slenderly lanceolate, flexuose, subtly undulate, and golden leaves give O. lineare a distinctive appearance. The species is frequently fruiting, and the erect, weakly sulcate capsules atop slender setae provide a further means of recognition. When dry, the mature exostome teeth are characteristically strongly incurved and erect endostome segments are conspicuous.
When sterile O. lineare could be confused with Leptobryum pyriforme or Pohlia tenuifolia. The Orthodontium lacks the red axillary hairs that characterise L. pyriforme. Leptobryum has more clearly differentiated oblong leaf bases and setaceous subulae with denticulate leaf apices. Pohlia tenuifolia has much shorter leaves than O. lineare (mostly 1.8–3.2 mm vs 3–6 mm) and is dioicous. When fruiting, Orthodontium is distinct from both these species by its erect, sulcate capsules, and the absence of an endostomal membrane. Confusion sometimes occurs with Campylopodium lineare. Orthodontium lineare differs from C. lineare by having a double peristome, longer and flexuose rather than distinctly cygneous setae, longer capsules, and numerous other less easily observed characters.
NI: N Auckland (Bay of Islands), S Auckland, Gisborne, Hawke’s Bay, Wellington; SI: Nelson, Marlborough, Canterbury, Westland (Craigieburn pākihi), Otago (Pine Hill, Dunedin region, Tahakopa Bay), Southland; St; Ch (Pitt I.); A; C. Reported from M by Seppelt (2004).
Austral. Tasmania*, mainland Australia*, South Africa*. Reported from southern South America and widespread and adventive in Europe (Smith 2004).
Epiphytic, on rotten wood, or terrestrial. When terrestrial, occurring mostly on humic or peaty soil, and less often on mineral soil. Tree-fern trunks, bases, and stumps provide the most common substrate, but O. lineare can also occur on accumulated humus on a variety of native and introduced trees including species of southern beech and the introduced conifers Pseudotsuga menziesii and Sequoia sempervirens. Also occurring on well-rotted, usually dry wood (often beneath overhangs) in a wide range of forest types and on thermally heated soil. Although O. lineare is widespread on both main islands, it appears to be a very uncommon plant in N Auckland and Westland L.D. (from the latter it is known only from a single depauperate collection, P. Beveridge s.n., 30 Oct. 2000, WELT M033415) and no collections have been seen from Taranaki L.D. The uncommonness of O. lineare in western parts of the main islands is mirrored by its relative rarity in the western parts of Britain (Smith 2004), where it is adventive. On the North I. ranging from near sea level (Mana I., Wellington L.D.) to c. 1100 m (Whakapapa, Wellington L.D.) and on the South I. and Stewart I. / Rakiura ranging from very close to sea level (Tahakopa Bay, Otago L.D.) to at least 900 m elevation (Mt Robert, Nelson L.D.). Campylopodium lineare, Leptotheca gaudichaudii, and Rhizogonium distichum are frequent associates, and Calomnion complanatum often co-occurs in epiphytic habitats.
Masses of modified (reduced, pale, and falcate) leaves occasionally develop on sterile plants and serve an asexual reproductive role. When growing on thermally heated soil the stems can become quite elongate (to c. 40 mm) and branched (e.g. K.W. Allison 516 from Rotorua, S Auckland L.D., CHR 490367).
Given the range of variation of O. lineare in N.Z., the features used by Meijer (1952) to distinguish O. ruahinense are unconvincing. The paroicous and gonioautoicous sexuality (of Meijer’s type) is representative of O. lineare. While more than one seta per perichaetium has been observed only in Meijer’s type, more than one fertilised archegonium per perichaetium has been observed in other N.Z. collections (e.g., K.W. Allison 3025, CHR 490370). The seta length (mostly 5–7 mm) and leaf length in Meijer’s type fall at the lower and upper end, respectively, of the range of continuous variation of N.Z. O. lineare. The lack of peristome tooth papillae cited by Meijer seems an unreliable feature as all capsules in the type of O. ruahinense are old and imperfect. The weight of evidence suggests that O. ruahinense does not deserve taxonomic recognition.
Shaw (2012) treated Orthodontium for Australia and recognised two species, O. lineare Schwägr. and O. pallens (Hook.f. & Wilson) Broth. [Nat. Pflanzenfam. 1, 3: 544, 1903]. He considered the latter species to be “possibly endemic to Australia” and distinguished it from O. lineare by a smaller stature and a more rudimentary endostome. Shaw also placed four Tasmanian or mainland Australia species into synonymy with O. lineare Schwägr.
Orthodontium lineare is an example of a southern hemisphere moss that has been introduced into Europe (probably from South Africa). It was first recorded from England by Watson (1922), who described it as a variety of O. gracile. The expanding range of O. lineare in Europe has been documented by Meijer (1952) and Ochyra (1982).