Class
Family
Genus
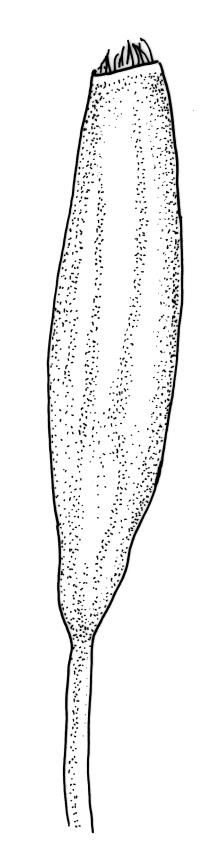
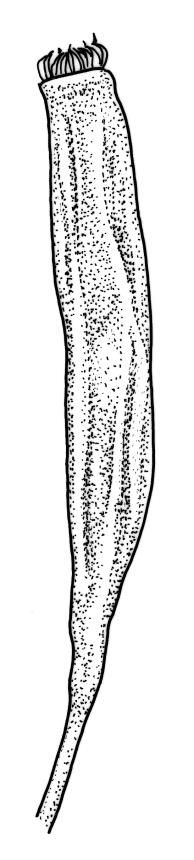
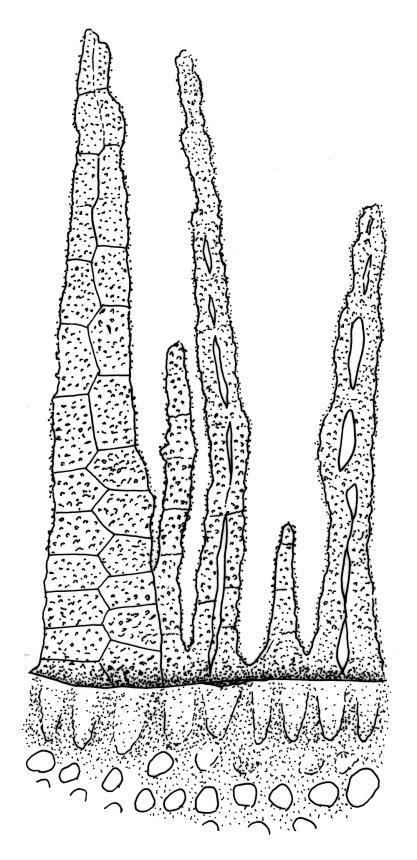
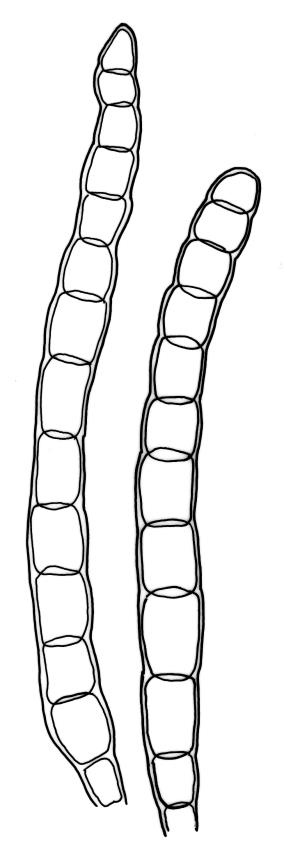
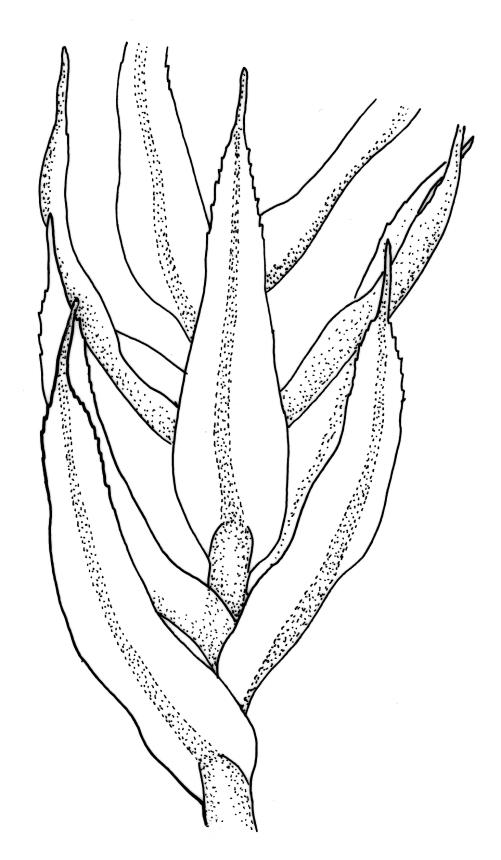
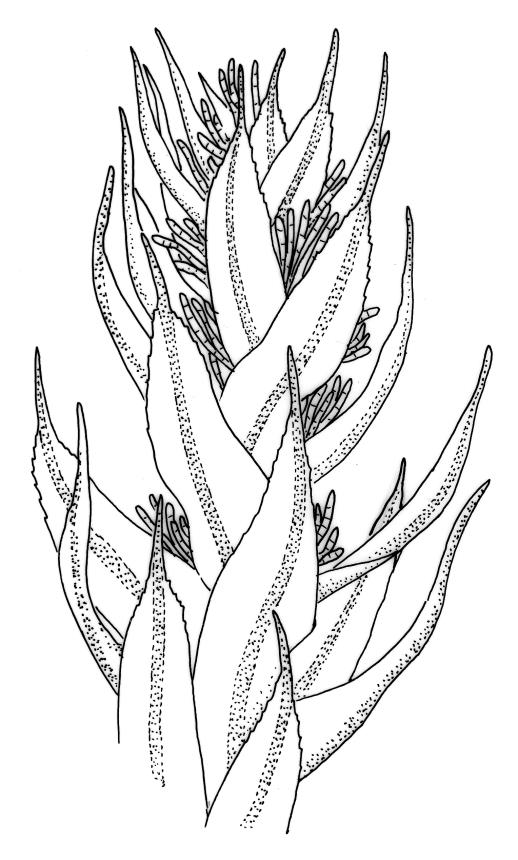
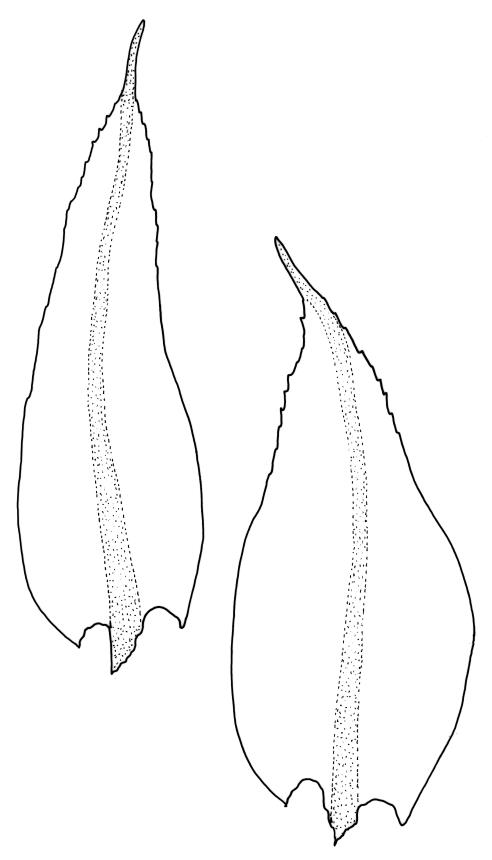
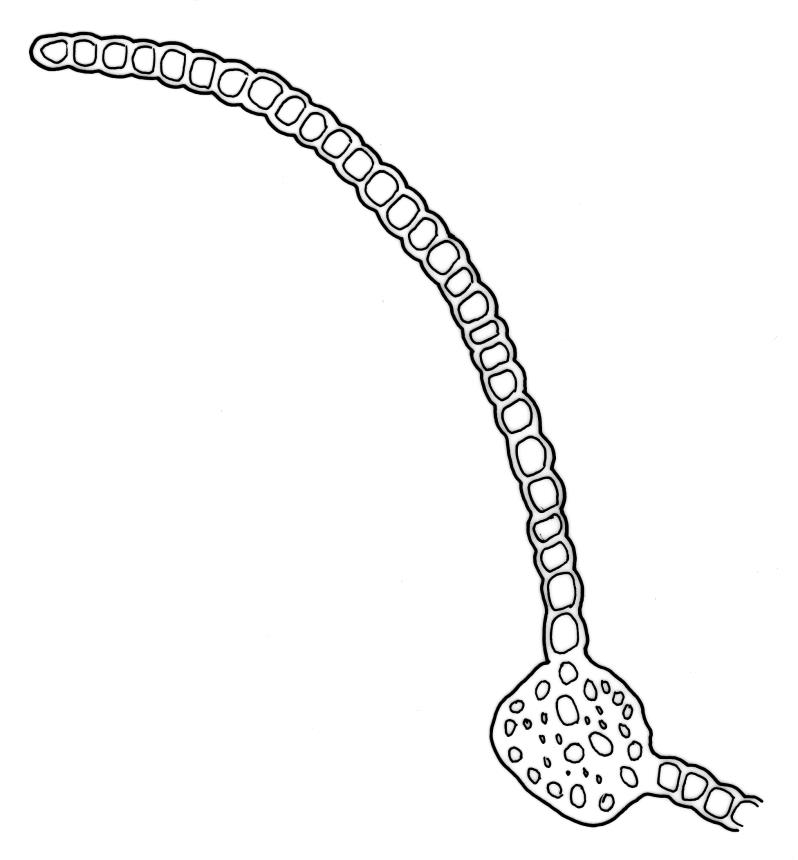
Plants rather small to medium-sized, slender, yellow-green to dark green, forming turves or dense cushions. Stems erect, branching by both forking and subperichaetial innovation (or occasionally with innovations arising within the perichaetium), <5 to c. 50 mm, in cross-section with 2–3 outer layers of thick-walled and pigmented cells and a distinct central strand, beset below with dimorphic red-brown rhizoids (the finer smooth, the more robust densely but finely papillose). Leaves spreading when moist, more erect and curved inwardly when dry, arranged in several rows but sometimes ± complanate near the stem apices or in small plants, ovate-lanceolate to linear-lanceolate (occasionally broadly ovate, especially on upper stems or innovations), aristate, weakly concave, plane at margins, not bordered, symmetric, narrowly decurrent, ± coarsely and irregularly toothed above; c. 1.6–2.2(–3.1) × 0.5–0.8 mm and c. 2.4–4(–5):1, strongly reduced in size near stem base and ± reduced near stem apex; upper and lower laminal cells ± isodiametric, incrassate, smooth, mostly 9–14 µm in greater diam.; marginal cells and alar cells not differentiated. Costa stout, ± sinuose, c. 60–75 µm wide in lower leaf, stoutly and short excurrent as an arista, smooth abaxially, in cross-section rounded and protruding abaxially, with median guide cells and both abaxial and adaxial stereid bands, a single abaxial cell layer ± enlarged. Gemmae mostly present and abundant in upper leaf axils of sterile plants, conspicuous (under hand-lens), linear, orange-brown to nearly black, transversely septate and uniseriate, (5–)8–15 cells and (120–)240–410 µm long.
Dioicous. Perichaetia terminal, usually overtopped by innovations, often serially; the leaves narrower and longer than vegetative leaves and often ± secund. Perigonia terminal and usually obscured by innovations. Setae straight, slender, pale brown, twisted weakly to the right above, mostly c. 15–25mm; capsules erect, straight or weakly curved, narrowly cylindric, 8-ribbed, mostly 3.0–4.5 mm, with a weak neck; exothecial cells oblong and firm-walled; annulus of 2–3 rows of inflated and hyaline cells, mostly falling with operculum; operculum bluntly conic, not rostrate. Peristome double; exostome teeth c. 300 µm, obtuse at apex, abaxially papillose throughout, with a divisural line becoming nearly straight in upper portions, adaxially surface papillose and transversely barred but not or scarcely trabeculate; endostome pale, with a very low membrane, the segments nearly equal the teeth, papillose and narrowly perforate throughout, alternating with single, variably developed, and sometimes nodose cilia. Calyptra cylindric and enclosing the entire capsule, splitting on one side or not, smooth, falling early. Spores nearly smooth, mostly 10–14 µm.
Brotherus 1924, fig. 387; Sainsbury 1955, pl. 41, fig. 2; Churchill & Buck 1982, figs. 3–4, 14–16; Malcolm & Malcolm 2003, p. 40; Meagher & Fuhrer 2003, p. 112; Seppelt et al. 2019, pl. 77.
Leptotheca gaudichaudii is most likely to be confused with Pyrrhobryum mnioides subsp. contortum and P. bifarium. From both these species, L. gaudichaudii differs by having unpaired marginal teeth and short basal laminal cells. The abaxial costal surface in L. gaudichaudii is smooth or inconspicuously toothed, while those in Pyrrhobryum spp. are conspicuously toothed.
When sterile L. gaudichaudii could easily be confused in the field with species of Pohlia, but the very compact, isodiametric cells (which can be observed with a hand-lens) distinguish the present species.
NI: N Auckland (Te Paki) including offshore islands (LB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; A. Reported from C by Vitt (1974).
Austral. Tasmania*, mainland Australia*, temperate South America*, South Georgia*. Reported from Falkland Is and South Africa by Churchill & Buck (1982). Adventive in Ireland*.
A common and widespread species occurring in a wide range of habitats, but most abundant in terrestrial habitats in southern beech forest, where it often occurs on exposed roots, but is also common on rotten wood, earth mounds, soil banks, and humic and mineral soil over rock, including greywacke, granite, and limestone. Often on tree-fern and tree trunks (especially southern beech). Occurring in a wide range of native forest types, scrublands, and grasslands, including those dominated by Chionochloa spp., and in exotic tree plantations. Leptotheca gaudichaudii occasionally becomes detached from the substrate to form "moss balls". Not seen north of Mt Ruapehu (J. Beever, pers. comm., 4 Aug. 2015) and there are no herbarium records of L. gaudichaudii from either the Waitakere Ranges (N Auckland L.D.) or the Coromandel Peninsula (S Auckland L.D.). The only record from N Auckland L.D. is a J.K. Bartlett collection from Te Paki Bush Scenic Reserve (WELT M008429), confirmed by Beever. Herbarium records suggest it is a common species in the Rotorua area, Gisborne, and the Mt Taranaki area. On the South I. it is a very common to abundant species in the St Arnaud Range and adjacent parts of the Nelson Lakes N.P., in the southern beech forest of the ranges of Canterbury and Otago, and in Fiordland N.P.
On the North I. ranging from c. 150 to 1220 m (Ruahine Range, Wellington L.D.) and on the South I. from near sea level (Milford Sound, Southland L.D.) to at least 1675 m (Spenser Range, Nelson L.D.). Its South I. distribution exhibits an eastern bias and it is known from relatively few localities west of the Main Divide. Frequently associated mosses include Bartramia papillata, Dicranoloma spp., Ditrichum cylindricarpum, Leucobryum javense, Pyrrhobryum mnioides subsp. contortum, Rhizogonium distichum, the hepatics Bazzania spp. and Lepidozia spp., and the lichen Cladia aggregata. When epiphytic, L. gaudichaudii often occurs with Leptostomum inclinans.
The presence of the orange-brown to nearly black filamentous gemmae is highly diagnostic and a useful field character. However, gemmae are not produced on fertile plants and are sometimes absent even from sterile plants. Perigonia are usually obscured by subtending innovations and are difficult to find, even in fruiting populations (e.g., A.J. Fife 5033a from Mt Owen, Nelson L.D., CHR 103615). Although individual stems in N.Z. specimens have leaves that are linear-lanceolate in shape, the leaves in the bulk of N.Z. material are ovate-lanceolate.
Some collections, particularly from higher elevations, have upper portions of the shoots decidedly complanate (e.g., A.J. Fife 6351 from Morgan Tarn, Nelson L.D., CHR 461742), but the distinction is not worthy of taxonomic recognition.
A Tasmanian endemic, L. wattsii Cardot, was reduced to varietal status by Churchill & Buck (1982), who noted that the material they had seen, including the Mt. Wellington type material collected by Watts, was intermixed with typical L. gaudichaudii. Type material of this name has been viewed only on JSTOR (image viewed online, JSTOR Global Plants, accessed 5 Aug. 2015.) and it apparently consists of only a few sterile shoots. No other material annotated by Churchill or Buck has been available for examination. However, L. gaudichaudii has variable leaf shape in N.Z., and in some N.Z. material leaves of upper shoots or innovations can be ovate, approaching those illustrated by Churchill & Buck for the var. wattsii. The Tasmanian taxon seems doubtfully worthy of recognition. If var. wattsii were to be accepted, then N.Z. material would, of necessity, be termed L. gaudichaudii Schwägr. var. gaudichaudii. However, it seems desirable to avoid this cumbersome name here given the doubtful status of the Tasmanian taxon.