Class
Family
Genus
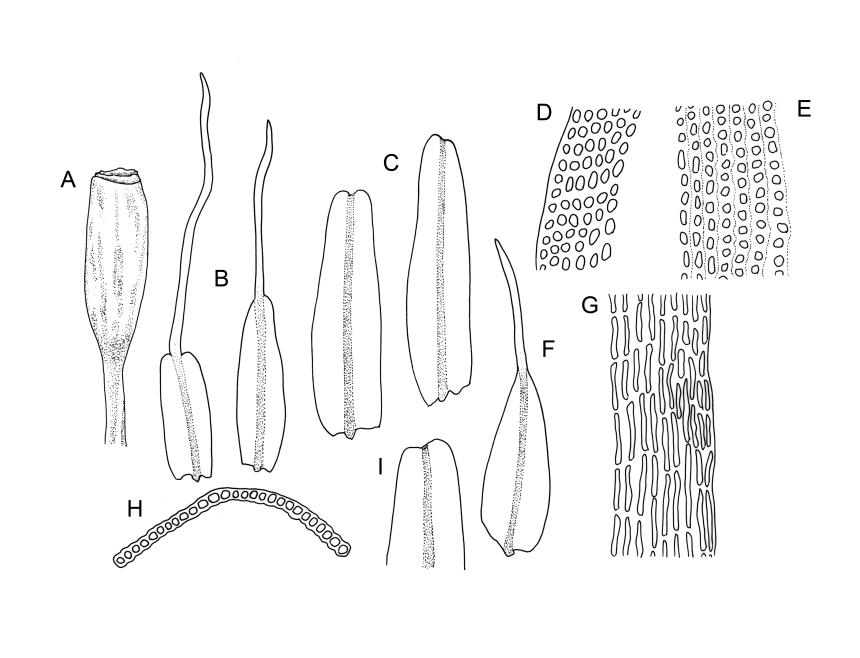
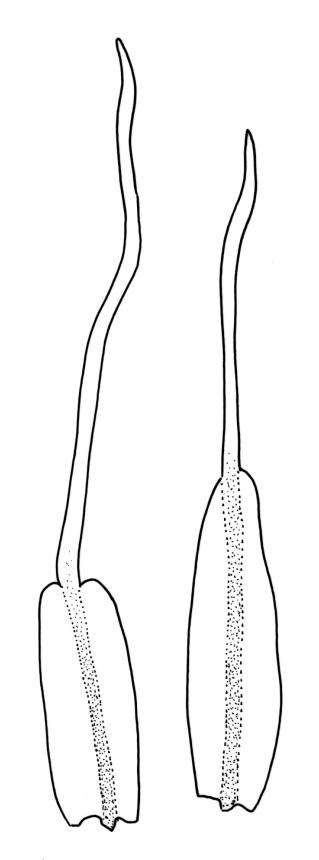
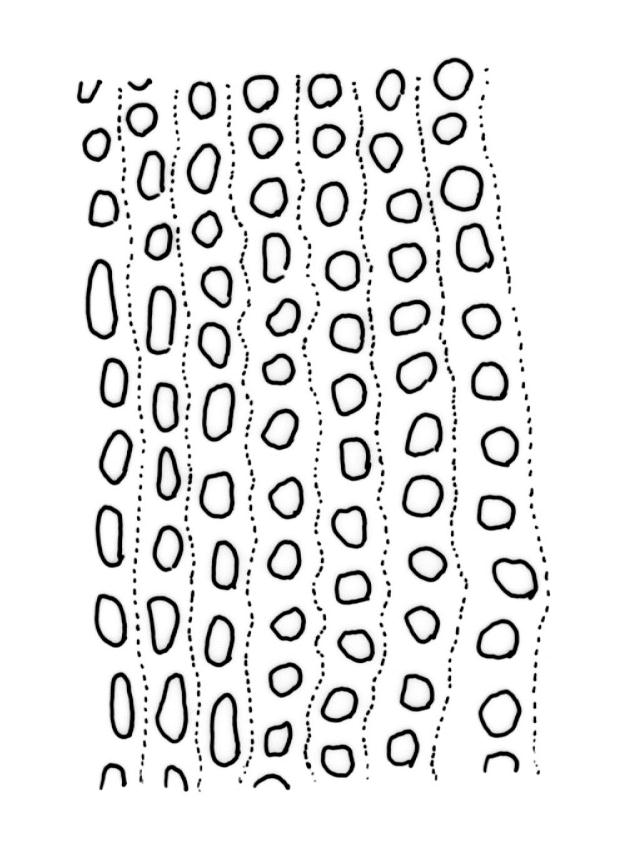
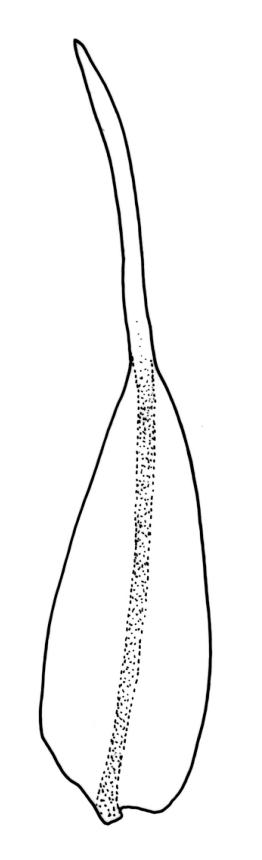
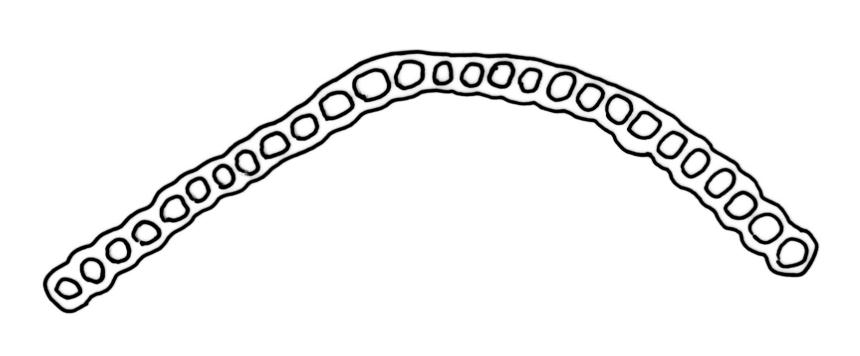
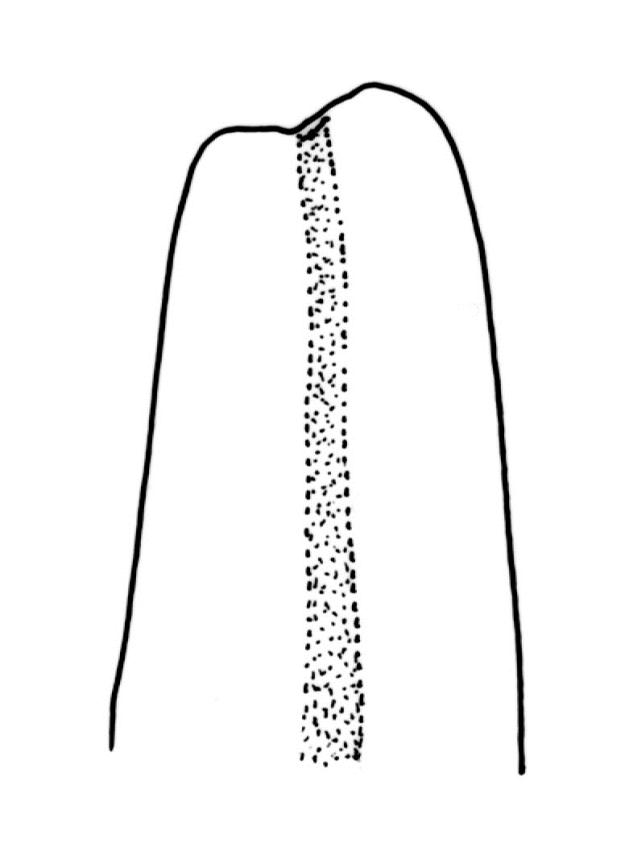
Plants slender, lustrous, bright-green to gold-green above, gold-brown below. Branches ± stiff, erect, mostly simple but forking below the perichaetium in ♀ plants, penicillate, c. 6–15(–20) mm. Stem leaves and lowermost branch leaves erect-flexuose when dry, lanceolate-acuminate, with the costa excurrent. Branch leaves curved-flexuose and spiralled (usually tightly) and some leaves reflexed when dry, erect-spreading and straight or slightly flexed, not funiculate when moist, oblong or ligulate, retuse, broadly obtuse (after loss of the arista) or sometimes irregular at apices, keeled, usually with a single plication below, young leaves with a long, linear, stiffly flexuose arista which usually breaks off before leaf maturity, c. 1.4–2.0(–2.7) mm (before loss of the arista), and 1.3–2.0 mm (after loss of the arista); margins plane, entire (except for apical notch); upper laminal cells rounded-quadrate to short rectangular-elliptic, clear, smooth, bulging, and very thick-walled, unistratose; mostly 7–12(–20) μm long, with lumina 6–13 × 6–7 μm (slightly smaller at margins); mid laminal cells mostly similar to upper; in distinct longitudinal rows; inner basal cells mostly elongate, irregularly thick-walled, with straight lumina, smooth, plane or slightly bulging; 16–27(–40) μm long (with lumina c. 2–4 μm wide); basal marginal cells not forming a distinct border. Costa long-excurrent to form an arista commonly extending c. 700–1300 μm beyond the lamina of immature leaves, the abaxial superficial cells elongate throughout; in cross-section with 1–2 layers of c. 4–6 guide cells, c. 2–3 layers of abaxial stereids, and 1–2 layers of adaxial stereids. Gemmae absent.
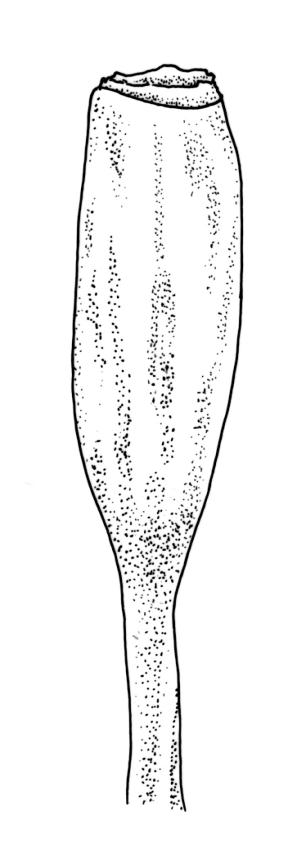
Dioicous or pseudautoicous (male plants not seen). Perichaetial leaves smaller than adjacent vegetative leaves, ovate-elliptic, with fragile aristae, 1.3–1.5 mm (after fall of the arista). Perigonia not seen. Setae 4.0–7.0 mm, smooth, straight or slightly flexuose, thick, dextrorse; capsules narrowly ovate to fusiform-cylindric, not constricted at mouth, indistinctly ribbed to almost smooth, ± erect, c. 2.0 mm; exothecial cells thick-walled, elongate-sinuose to elliptic, apparently shorter and thicker-walled on ribs, variable in length (to c. 65 μm), becoming isodiametric near rim. Peristome probably double; exostome teeth 16, recurved when dry, incurved when moist, warty-granulose, linear-lanceolate, often very thick outwardly and ± smooth below; endostome not seen. Calyptra deeply lacerate, smooth, strongly plicate. Spores not seen.
Sainsbury 1955, pl. 37, fig. 1 (as Macromitrium longirostre var. caducipilum); Vitt 1983, figs 34–43, 45–46.
Macromitrium retusum and M. helmsii are superficially similar and have been confused; both have penicillate branches, with the immature leaves abruptly contracted to a long and fragile arista, and with mature leaves almost always retuse. However, M. retusum is made distinct by its smooth and clear upper laminal cells, and branch leaves tightly spirally twisted around the branch (but with individual leaves often curved outward) when dry. By contrast, the branch leaves in M. helmsii have upper laminal cells with dense and multiple papillae and are "individually incurved-twisted with the upper portions curved outward and down when dry". I concur with Vitt’s observation (1983, p. 46) that the leaves of M. retusum are more ligulate to oblong and usually not much wider in the lower portion, whereas those of M. helmsii are more lanceolate, widest below, and gradually narrowed upward. Vitt (1983, p. 23) clarified the previously confused nomenclature of M. retusum and M. helmsii.
NI: N Auckland (Maunganui Bluff, near Auckland City), S Auckland (Mt Karioi), Gisborne (Māhia Peninsula), Wellington (Tangiwai, Karehana Bay, Tītahi Bay, Wilton’s Bush); SI: Nelson (Mt Burnett, Kōhaihai River, Punakaikī), Marlborough (s. loc.), Westland (Greymouth, Pāroa, Haast), Canterbury (near Christchurch), Otago, Southland; St; C. The Māhia Peninsula and Christchurch records are based solely on Vitt (1983).
Endemic.
Found largely on coastal tree trunks and branches and on coastal rocks. It is rarely collected on the North I. but is frequent in coastal areas in the southern part of the South I., particularly from Dunedin south and west to at least the vicinity of Riverton (Southland L.D.). It appears to be scattered (or poorly collected) in Westland. Vitt (1983, fig. 51) annotated a small number of Westland specimens but did not include these in his distribution maps, and there are a small number of more recent Westland collections. It is common on Stewart I. (where W. Martin, in herb., termed it "probably the commonest species" [of Macromitrium] in the north portion of that island. Vitt (1983) considered it uncommon on Campbell I., where he stated it to be found only on Dracophyllum in dense riparian forests, usually in association with M. longirostre. It occasionally occurs in inland situations on both main islands (e.g., Mt Karioi, Tangiwai, the Horse Range (Otago L.D.), and Lake Hauroko (Southland L.D.)). While primarily occurring at low elevation coastal sites, this species has been collected at elevations as high as 641 m on the South I. (Mt Burnett) and 560 m on the North I. (Mt Karioi).
Host woody species include Brachyglottis repanda, Carpodetus serratus, Coprosma linariifolia, C. rotundifolia, Fuchsia excorticata, Melicytus ramiflorus, and Brachyglottis rotundifolia, as well as the podocarps Dacrydium cupressinum and Podocarpus totara. It also occurs on coastal granite and dolomite. Frequently associated mosses include Calyptopogon mnioides, Macromitrium gracile, M. longipes, M. longirostre, and Orthorrhynchium elegans.
My observations on perichaetial leaves (seen most clearly in W. Martin 289.5 from Riverton, Southland L.D., CHR 629455 and W. Martin s.n. 25.1.49 from Stewart I., CHR 629454) differ from Vitt’s and suggest they are smaller and more ovate than the adjacent vegetative leaves, measuring c. 1.3–1.5 mm after the arista has fallen.
Vitt (1983) considered M. retusum to be "structurally stenotypic" albeit with "some variation of leaf set and basal cell length". According to Vitt: "the amount of individual twisting of branch leaves is variable, with some specimens possessing uniformly non-twisted, spirally curved leaves and others having a variable number of leaves curved outward as well as spirally positioned around the branch. The basal leaf cells exhibit similar variation as those of M. longirostre. Most populations have elongate basal cells with only a few rows of shorter mid leaf cells extending into the basal area, particularly near the costa. However, a few populations have short (rounded-elliptic to shortly rectangular) cells extending nearly to the leaf base, and the more typical elongate basal cells are limited to the insertion, basal margins, or interspersed among the short cells". He observed a range of basal cell shapes in single plants and did not consider this feature to be taxonomically significant.
Vitt & Ramsay (1985) defined a "Macromitrium longirostre group" that included M. retusum, M. longirostre, and M. ramsayae (recognised at the varietal level here). The fragile leaf tips and retuse mature leaves distinguish M. retusum from these closely related taxa.