Class
Family
Genus
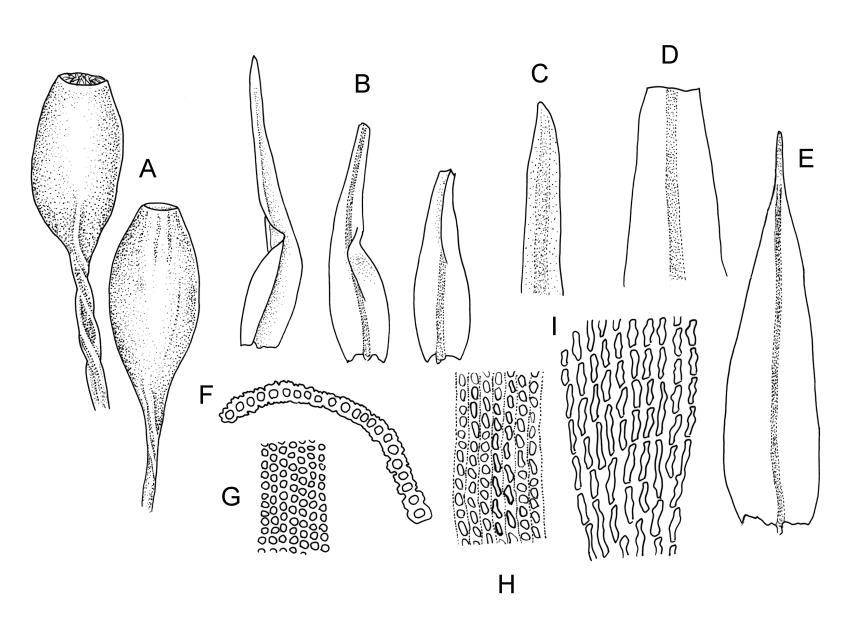
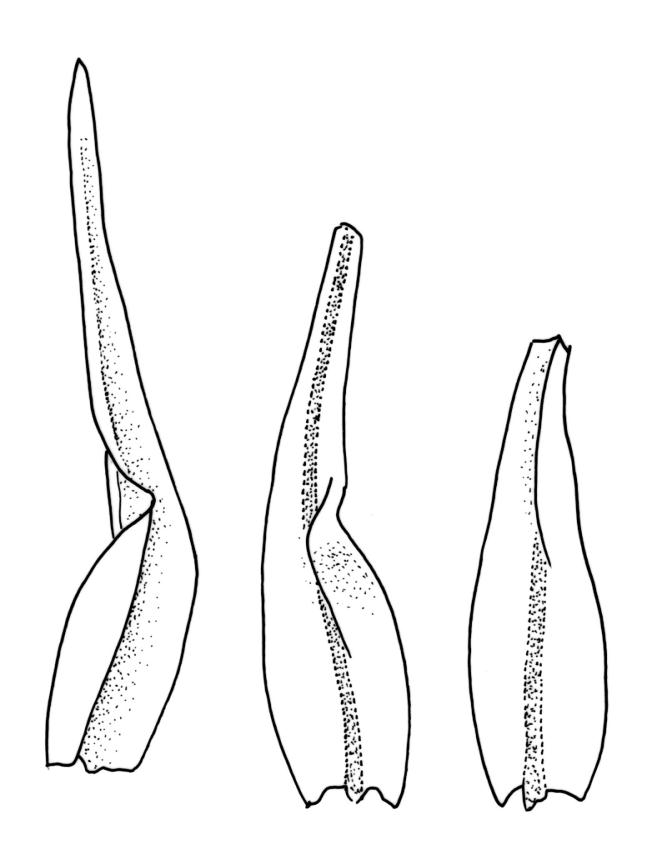
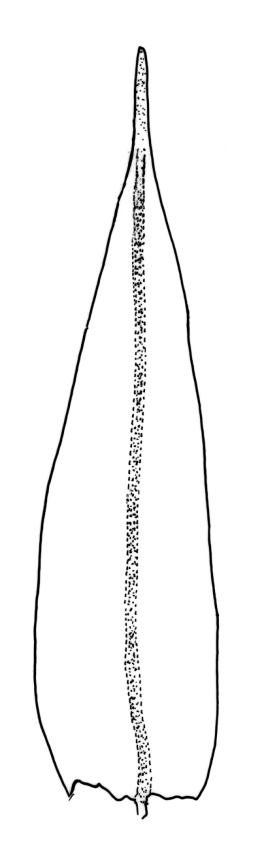
Plants medium-sized to moderately robust, slender, stiff, not lustrous, usually light green, yellow-green, or bronze above, olive-green or brown below. Branches usually 5–20(–40) mm, erect, stiff, often once-branched by subperichaetial innovation. Stem leaves erect-flexuose when dry, spreading-recurved moist, ovate-lanceolate, 0.8–1.0 mm. Branch leaves precisely twisted, indistinctly funiculate, and with upper portions incurved (usually to one side) or decurved when dry, ± wide-spreading-flexuose and clearly funiculate when moist, lanceolate from an oblong or ovate base, gradually narrowed to a subulate or ligulate apex, keeled, frequently with apices broken off below but with some intact near branch apices, 1.8–2.8 mm (when intact); margins entire, broadly reflexed at mid leaf; upper laminal cells ± quadrate, thick-walled, ± bulging, densely pluripapillose and often obscure, arranged in indistinct longitudinal rows, 6–8(–10) µm wide, sometimes becoming larger (to c. 15 µm in greater dimension), more irregular, and ± bistratose in subula; mid laminal cells slightly larger and more distinctly ranked, transitioning gradually with the inner basal cells; inner basal cells elongate, thick-walled, nearly smooth or tuberculate, mostly 20–50 × 8–10 µm (lumina c. 2–3 m wide; curved or nearly straight); marginal basal cells longer, but not forming a distinct border. Costa ending in or just below apex (in intact leaves), the abaxial superficial cells elongate and smooth, in cross-section with one adaxial row of c. 4 guide cells and 2–3 rows of abaxial stereids. Gemmae absent.
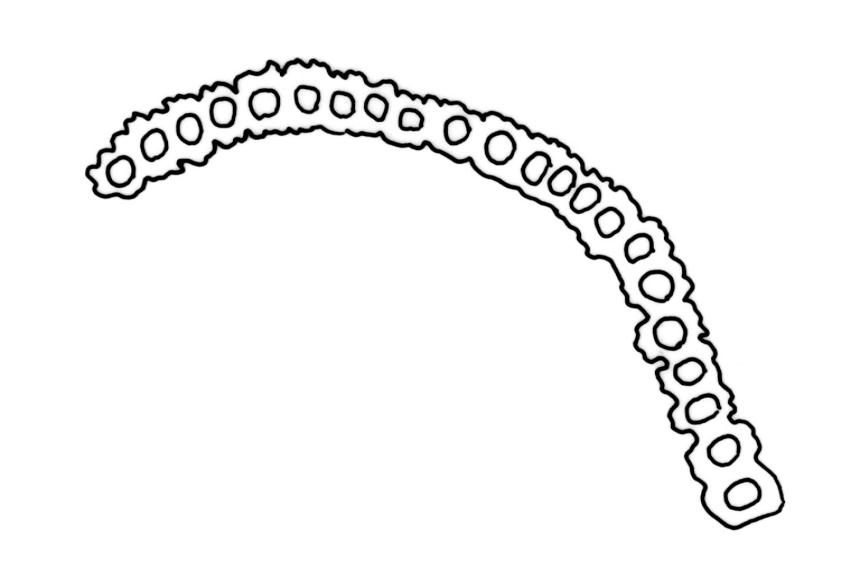
Pseudautoicous. Perichaetial leaves clearly differentiated and mostly distinctly larger than vegetative, erect and sheathing, lanceolate to oblong-lanceolate, usually with intact apices, 2.2–3.0 mm. Dwarf male plants epiphyllic, with stems to c. 1 mm and sometimes with multiple (to 3) perigonia, the perigonial bracts spreading. Setae 2.5–7 mm, smooth, ± straight or moderately flexuose, sinistrorse; capsules 1.0–1.7(–2.2) mm, ovate, (becoming more broadly ovate with age), smooth or weakly 8-sulcate; exothecial cells short and rather irregular, elliptic to rhombic-rectangular, thick-walled, mostly 20–40 µm long sometimes with bands of longer (30–60 × c. 6–8 µm) cells, shorter and thicker-walled at rim; stomata and annulus as per genus; operculum erect, c. 1 mm. Peristome single; exostome teeth 16, well-developed, inflexed-erect when dry, pale to white, c. 200 µm, coarsely papillose on both surfaces, often fragile with age. Calyptra c. 3 mm, deeply and evenly lacerate, shiny, golden, strongly plicate, naked or nearly so. Spores indistinctly anisosporous, 16–40, finely papillose.
The upper lamina forms a long, flattened bistratose subula, which is fragile in many plants but, when intact, becomes conspicuously decurved giving a distinct appearance to the plants. Often the intact leaves are restricted to the branch tips and this combines with the other features cited above to give the plants a distinctive habit. I agree with Vitt (1983) that populations with extended fragile leaf apices and those with acute, mostly non-fragile apices grow together and do not appear to be differently distributed.
Specimens with short, intact apices sometimes occur and these can be confused with M. microstomum or M. prorepens. Such material of M. gracile is distinct by having truly lanceolate leaves, obscure and pluripapillose upper leaf cells, stiff branches, and single innovative branches below the perichaetia.
Confusion sometimes occurs with the related M. helmsii. However, in forms of M. gracile with protracted leaf apices, these are present in at least some fully mature leaves, gradually tapered to the apex, and decurved. In M. helmsii, by contrast, the protracted leaf apices extend beyond the stem apex in the same axis as the stem "in apical tufts", and (as explained well by Vitt 1983, p. 47) the aristae reach their "mature length first, and then lamina development [takes] place secondarily. Thus mature aristae [terminate] short immature leaf laminae and once the laminae reach full length, the aristae are broken off." The irregular nature of the broken leaf edge in M. gracile contrasts with the retuse leaf edge in M. helmsii. These features, once seen, are easily recognised.
Macromitrium gracile is readily distinguished from M. longipes, which also has elongate perichaetial leaves, by plants being dull in appearance (due to the papillose laminal cells), the less strongly funiculate and frequently broken leaves, and the strongly sheathing perichaetial leaves. By contrast, M. longipes is lustrous (due to smooth laminal cells), more strongly funiculate (readily seen in dried material), and has unbroken leaves and less sheathing perichaetial leaves.
Vitt & Ramsay’s (1985) "M. gracile group" includes five species in Australasia. Macromitrium gracile and M. helmsii are N.Z. endemics. Macromitrium incurvifolium is predominantly distributed in the tropical Pacific and eastern Asia and occurs in the Kermadec Is.
K; NI: N Auckland, including offshore islands (TK, PK, HC, LB, GB, RT), S Auckland, Gisborne (Lake Waikaremoana), Hawke’s Bay, Taranaki (Mt Taranaki), Wellington; SI: Nelson, Marlborough, Canterbury (Arthur’s Pass, Banks Peninsula), Westland, Otago (Mt Cargill, Tautuku), Southland; St; Ch.
Endemic.
Macromitrium gracile is particularly frequent on Carpodetus serratus, but it occurs on a wide variety of native flowering tree species including Aristotelia serrata, Coprosma spp., Fuchsia excorticata, Griselinia lucida, Lophomyrtus bullata, Melicytus ramiflorus, Metrosideros excelsa, M. umbellata, Neomyrtus pedunculata, Olearia ilicifolia, Pseudowintera colorata, and Vitex lucens, as well as the native gymnosperms Dacrydium cupressinum, Podocarpus totara, Prumnopitys ferruginea, and P. taxifolia, and the introduced genera Cupressus and Berberis. It occurs infrequently on southern beech, but it has been recorded from Fuscospora solandri s.l., F. fusca, and Lophozonia menziesii. On the Chatham Is this species is also recorded from Corynocarpus laevigatus, Dracophyllum arboreum, and Myrsine coxii. It rarely occurs on rock (as at D’Urville I., Marlborough L.D., J.E. Beever 46–58b, CHR 461786). Observations on recently fallen trees and branches suggest that M. gracile is more abundant in the forest canopy than it is on the lower trunks of forest trees.
Frequently associated mosses include Calyptopogon mnioides, Camptochaete arbuscula, Cyrtopus setosus, Dicranoloma menziesii, Holomitrium perichaetiale, Leptostomum inclinans, Lopidium concinnum, Macrocoma tenue, Macromitrium retusum, M. helmsii, M. longirostre, M. orthophyllum, M. longipes, Orthorrhynchium elegans, Papillaria crocea, Tetraphidopsis pusilla, and Trachyloma planifolium. A wide variety of hepatics including species of Frullania, Lepicolea, Plagiochila, Plagiochilion, Porella, and Radula also occur with it. On the North I. occurring from sea level (at Whangārei, N Auckland L.D.) to 950 m (Mt Ruapehu, Wellington L.D.) and on the South I. from sea level (Dusky Sound, Southland L.D.) to at least 550 m (at Mt Cargill, Otago L.D.).
Macromitrium gracile is found in all forested and scrub areas on the North I., but is more common at lower elevations. On the South I. M. gracile is a very common epiphyte in the Marlborough Sounds and at Pelorus Bridge in Marlborough L.D., at many localities in Westland L.D. including Lake Kaniere, Kellys Creek and the Ōtira Valley, and at Tautuku in the Catlins region of Otago. It is a common species on both Stewart and Chatham Is. It is a rare species east of the main divide on the South I. Vitt (1983, fig. 115) mapped an occurrence near Lake Pūkaki (Canterbury L.D.), but I have seen no specimen from there.
In the field and in its most common form, the usually long and stiff branches, the shape and set of the leaves (mostly turned to one side and incurved in their lower portions but apically decurved and indistinctly funiculate when dry), together with the often-produced and clearly enlarged and sheathing perichaetial leaves make this species easily recognisable. When plants are dry, the leaves often appear (because of their inrolled set) to be intact but when moist and widely spreading their truncated apices are easily seen.
I concur with Vitt (1983, p. 42) that M. gracile is a variable species, which "exists in several intergrading phenotypes". Paraphrasing Vitt (1983), all of these forms exhibit lanceolate leaves, stiff, wiry branches with branching usually single beneath perichaetia, a distinctive leaf set (with each leaf incurved or inrolled with apices curved to one side) and leaf insertion spirally arranged, obscure and densely pluripapillose upper leaf cells in longitudinal rows, elongate basal cells, sheathing and conspicuous perichaetial leaves, smooth setae sinistrorse, and ovate, non-puckered capsules with well-developed peristomes.