Class
Family
Genus
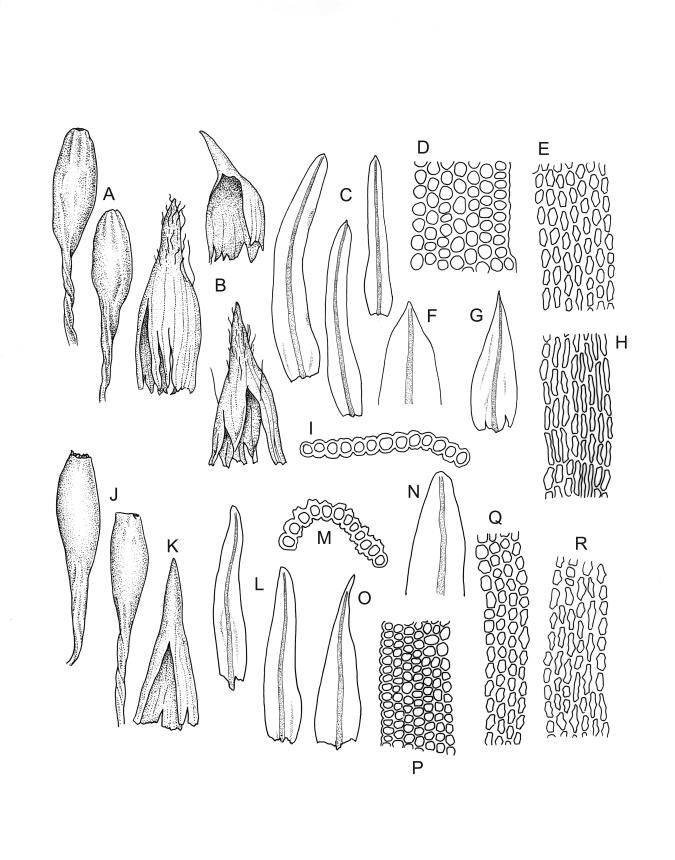
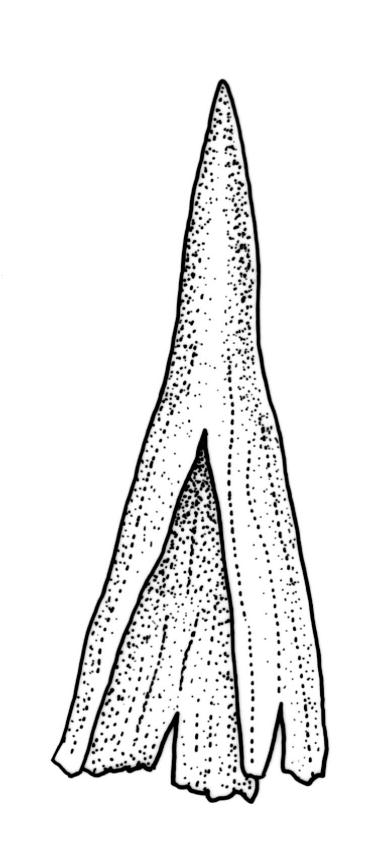
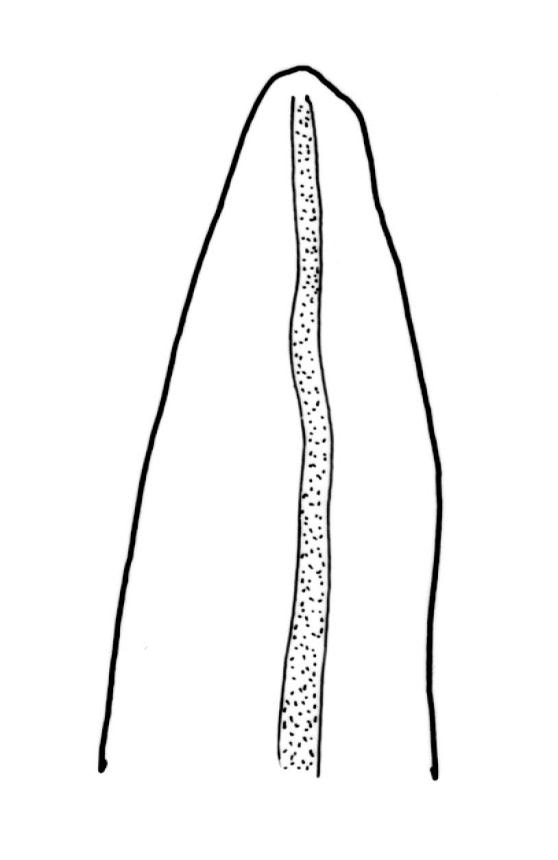
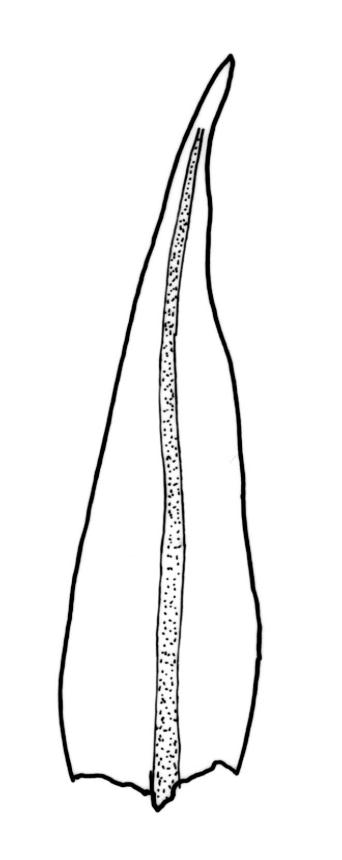
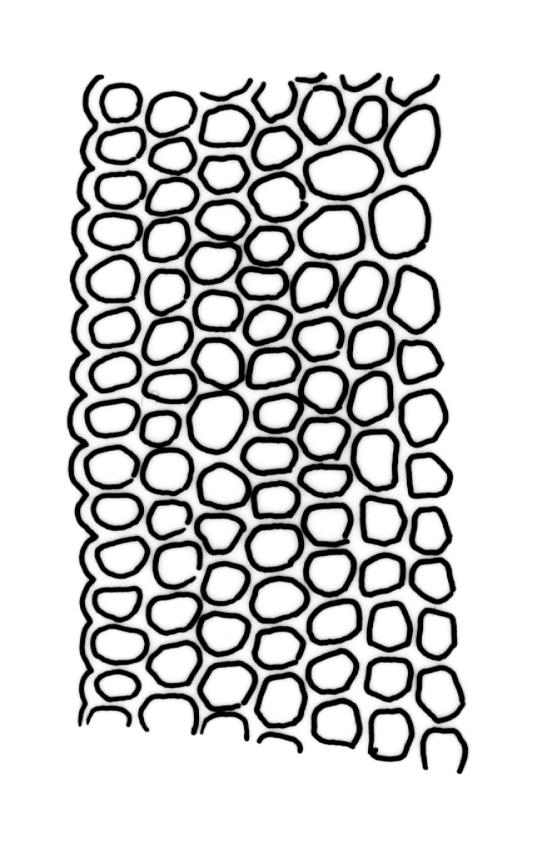
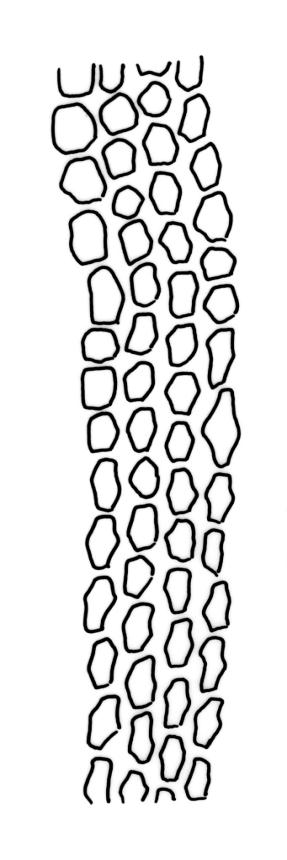
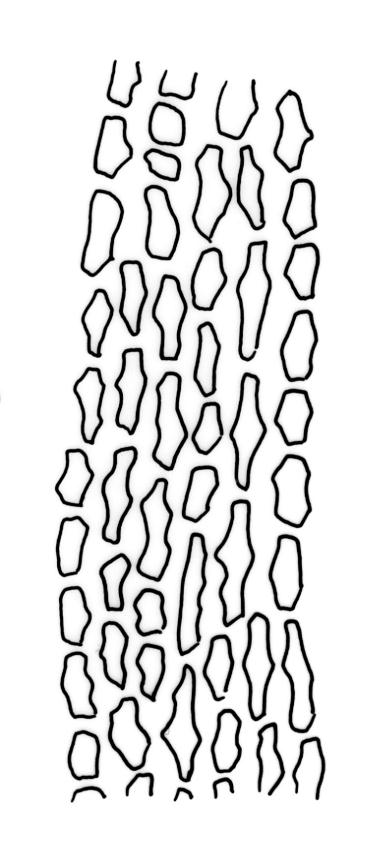
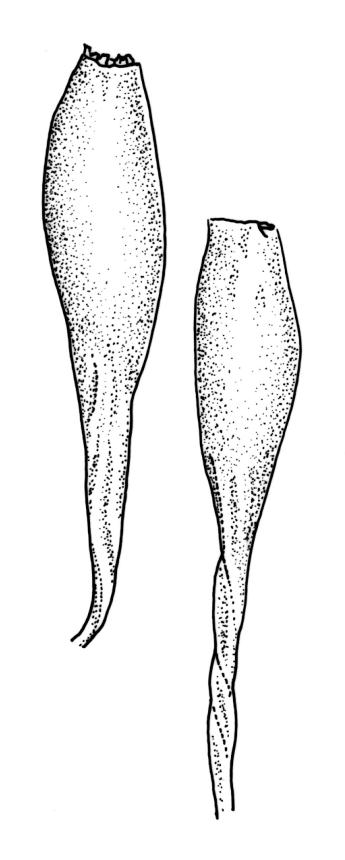
Plants slender, dull, yellow- to olive-green above, dark green or brown below. Branches to c. 17 mm, usually less. Stem leaves flexuose-erect dry, widely spreading and recurved when moist, lanceolate to broadly lanceolate, acuminate or acute, 1.0–1.3 mm. Branch leaves flexuose-twisted, with upper portion strongly inrolled and apices obscured when dry, flexuose-spreading with weakly inflexed apices when moist, not funiculate, ligulate to lanceolate-ligulate, rounded-obtuse to broadly acute, often with a 1-celled apiculus, strongly keeled, 1.5–2.5 mm; margins crenulate above, plane to ± reflexed and entire below; upper laminal cells isodiametric and rounded-subquadrate, thin-walled sometimes with weakly thickened corners, strongly bulging, usually with 1–4 small, conical papillae, unistratose, 9–15 µm wide, often smaller and transversely elliptic to rounded at margins; mid laminal cells similar, extending basally with little change far toward the leaf insertion (to within c. 0.1–0.25 of leaf length) and then transitioning abruptly into the inner basal cells; inner basal cells short-rectangular, 14–33 × c. 9 µm, (lumina c. 4–6 µm wide), ± thick-walled, porose, and mostly smooth; marginal basal cells not forming a distinct border. Costa ending a few cells below apex, shiny and smooth abaxially with elongate cells exposed (except near apex), in cross-section with 2–3 guide cells and 1–2 rows of abaxial stereids. Gemmae absent.
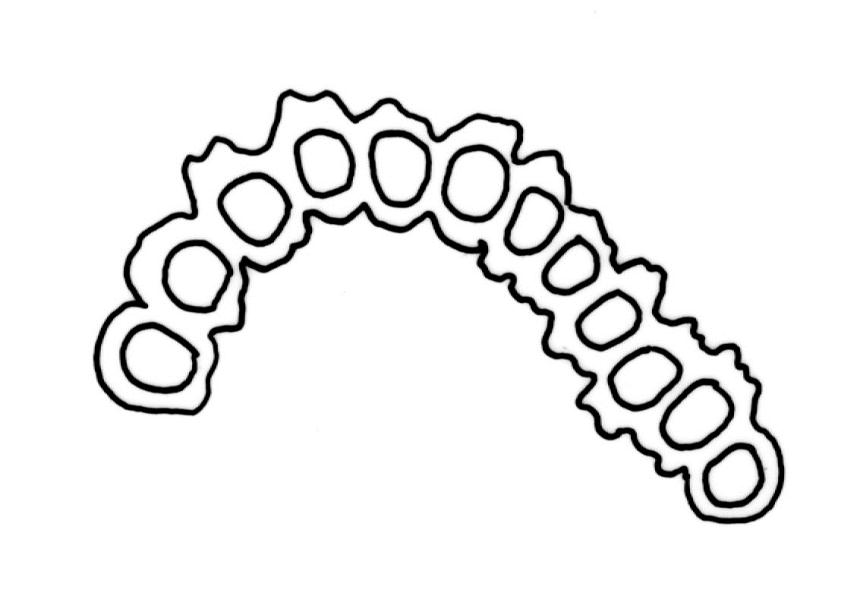
Pseudautoicous. Perichaetial leaves smaller than adjacent vegetative leaves, ovate-lanceolate, gradually acuminate or acute, not sheathing, 1.1–1.7 mm. Dwarf male plants axillary, bulbiform. Setae 2–8 mm, smooth, flexuose-erect, slender, sinistrorse; capsules narrowly oblong-ovate to cylindric-oblong, gradually contracted to the seta through a long, wrinkled neck, smooth, not plicate at mouth, usually with mouth darker and moderately constricted with age, c. 1.5–2.0 mm; exothecial cells thin-walled, ± elliptic-oblong to long-rectangular, 40–85 × 10–22 µm, more elongate and with thicker longitudinal walls at mouth; stomata and annulus as per genus; operculum erect-rostrate. Peristome single and apparently fugacious; exostome teeth 16, inflexed when dry, pale, blunt, transversely papillose-striate, c. 100 µm. Calyptra ± plicate, nearly entire or lobed at base, divided by 1–3 long slits, mitrate or approaching cucullate, not lacerate, smooth, naked. Spores anisosporous, 14–34 µm, finely papillose.
A capsule that is peristomate, yet collapsing at the mouth and without plications, is unique among N.Z. Macromitria. Closely related to M. ligulare is the very rare (in N.Z.) M. ligulaefolium, which can be distinguished, when fruiting, by having capsules strongly 8-plicate in the upper portion and having no peristome. I have not confirmed the differences in the exothecial cells (shortened above and not extensively longitudinally thickened in M. ligulaefolium) described by Vitt (1983). He also suggested that M. ligulaefolium could be distinguished by having an excurrent costa that in most leaves forms a slender apiculus. In my opinion, and that of J.E. Beever (pers. comm., 2012), these two species cannot be distinguished in the absence of well-developed capsules.
NI: N Auckland, including offshore islands (LB, GB), S Auckland, Gisborne, Hawke's Bay, Wellington; SI: Nelson (Nelson Lakes area including Travers Valley), Marlborough (Picton, Hāpuku River, Mt Fyffe), Canterbury, Westland (near Greymouth, Lake Kaniere, near Haast), Otago (eastern regions only); St.
Australasian. Mainland Australia*.
Vitt’s statement that this species is "infrequently collected north of Auckland" needs amendment, as there are numerous post-1983 collections from this region, mostly made by J.E. Beever. However, the paucity of material from near Mt Ruapehu and an apparent complete lack of records from Taranaki L.D. remain.
Macromitrium ligulare is most frequent in the lower-elevation podocarp-broadleaf forests of the central North I. Host species include Coprosma arborea, C. parviflora, Corynocarpus laevigatus, Leucopogon fasciculatus, Fuscospora solandri s.l., Knightia excelsa, Litsea calicaris, Myoporum laetum, Vitex lucens, and Podocarpus laetus. It also occasionally occurs on Cupressus macrocarpa and on rock (as at Trotters Gorge, Otago L.D). Bryological associates include Calyptopogon mnioides, Cryphaea sp., Leptodon smithii, Neckera hymenodonta, N. laevigata, Macrocoma tenue, Macromitrium gracile, Papillaria flexicaulis, and Zygodon hookeri. On the North I. ranging from near sea level in coastal forest (North Cape, N Auckland L.D.) to c. 900 m (Paeroa Range, S Auckland L.D.) and on the South I. from near sea level (Hāpuku River, Marlborough L.D.) to c. 850 m (Banks Peninsula, Canterbury L.D.).
When well-developed this is a distinctive species. According to Vitt (1983, p. 71) M. ligulare can be distinguished from other N.Z. Macromitria by a number of characters. These include: (1) "branch leaves [that] are inrolled with the apices hidden in the inrolled part and not curved to the side as in other species; (2) the strongly bulging, rather thin-walled upper leaf cells [that] are without large papillae and continue well below mid leaf and restrict the shortly elongate basal cells to the lower to ¼ (or less) of the leaf; (3) perichaetial leaves [that] are shorter than the vegetative leaves, but differ in shape; and (4) the completely smooth, delicate capsules that possess a peristome but that are collapsed at the mouth when mature. The exothecial cells are delicate and thin-walled in the body of the urn, but become thickened longitudinally at the rim, forming a distinct dark-brown rim associated with the collapsed area." The calyptrae are naked and split by one or two long slits. Vitt & Ramsay’s (1985) "M. ligulare group" includes the present species, M. ligulaefolium, M. incurvifolium, and two Australian endemics.