Class
Family
Minor elements in the following description are from Vitt (1980).
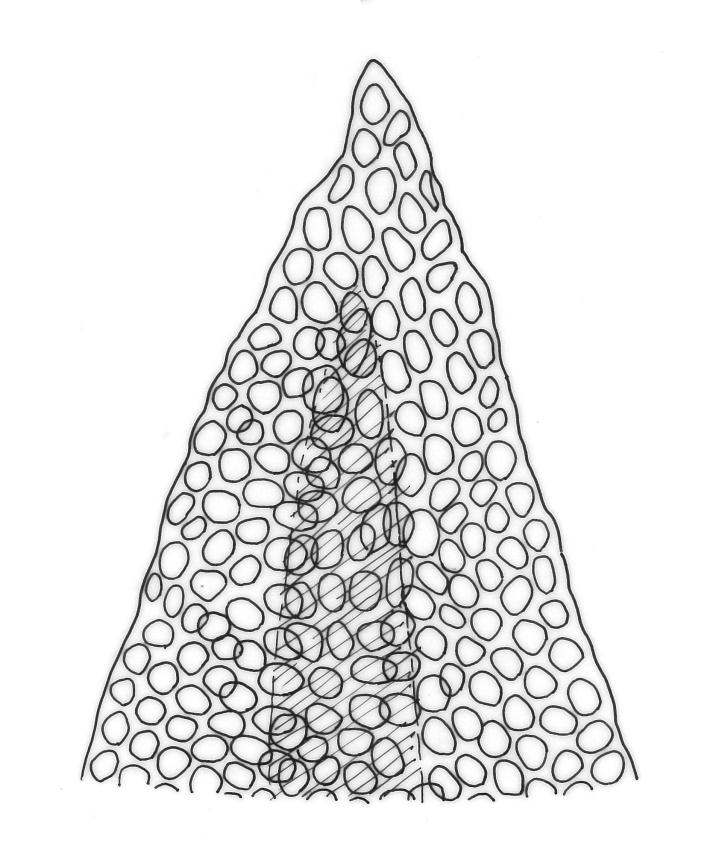
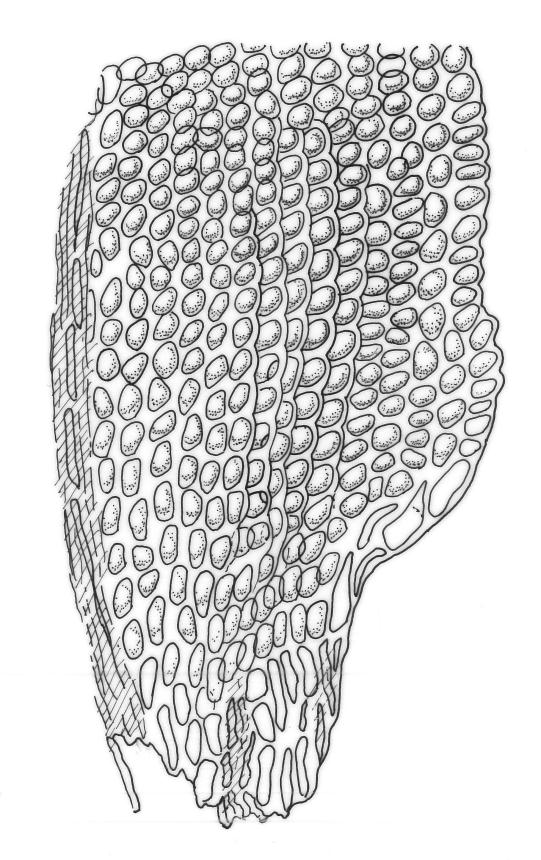
Plants gold-brown to dark green, forming loose and often extensive mats on bark. Stems creeping and elongate (c. 10 to >50 mm in N.Z. material), apparently lacking rhizoids, in cross-section with thick-walled outer cells and no central strand. Stem leaves weakly spreading when moist, appressed when dry, broadly ovate, acute, c. 1 mm. Branches weakly ascendant, mostly short (<7 mm), curved or straight when dry, simple or branching by innovation and forking. Branch leaves erect-imbricate or rarely squarrose when moist, not contorted when dry, oblong-ovate, acute (sometimes broadly), not fragile, weakly keeled above, strongly concave in the juxta-costal basal region, mostly 1.0–1.3 mm (but becoming progressively larger below perichaetia); margins entire and plane; upper laminal cells rounded, isodiametric to short-elliptic, firm-walled, not bulging, smooth, unistratose, c. 6–10 µm in greater dimension, becoming gradually strongly bulging and often ± oblate at mid leaf; inner basal laminal cells rounded-rhombic (never elongate), with a single strong and rounded projection but otherwise smooth, sometimes weakly nodose near the costa, mostly c. 12–18 × 8–10 µm; marginal basal cells usually more compact. Costa single, prominent, mostly fading in the apex or sometimes percurrent, the abaxial superficial cells in the upper half short and similar to laminal cells, in cross-section lacking guide cells and with cells ± uniform throughout. Gemmae absent.
Cladautoicous in N.Z. material. Perichaetial leaves enlarged (with the transition from vegetative leaves gradual). Perigonia gemmiform and often on short branches, readily found in fruiting material. Setae 3–6 mm, smooth, straight, weakly dextrorse or less often sinistrorse; capsules narrowly cylindric-ellipsoid, from a long but ill-defined neck, smooth throughout or becoming 8-sulcate below the mouth when dry and old, 1.5–2.0 mm; exothecial cells thick-walled, the longitudinal walls ± thicker than the transverse, mostly oblong and less than c. 45 µm, becoming smaller and forming indistinct bands of thicker-walled cells at the mouth; stomata superficial, restricted to the neck and lower portion of the urn; annulus not seen; operculum rostrate from a conic base, c. 0.8 mm. Peristome double, reduced to two low membranes, the exostomal membrane coarsely papillose. Calyptra large, plicate, densely hairy, enclosing the entire capsule and often slit on one side. Spores 1-celled, isosporous, round, thick-walled, 21–39 µm diam., papillose.
Scott & Stone 1976, p. 43 (as Macromitrium tenue); Meagher & Fuhrer 2003, p. 67 (as Macrocoma tenuis subsp. tenuis); Vitt et al. 1995, fig. 4 a–k.
In a N.Z. context, M. tenue could only be confused with a species of Macromitrium. However, it is usually easily recognised in the field by its epiphytic creeping stems with appressed and straight dry leaves and the frequently produced, large, and densely hairy calyptra enclosing the capsule on a relatively short seta. Additionally, the short, rounded-rhombic basal laminal cells (only the northern and epilithic Macromitrium brevicaule has basal laminal cells of a comparable shape) and a peristome reduced to two pale, low membranes provide further distinction. No species of N.Z. Macromitrium combines this unique suite of characters.
K; NI: N Auckland, including offshore islands (GB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington. SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland.
Anomalous. Eastern mainland Australia*. Reported from Tasmania and several countries of southern and eastern Africa by Vitt (1980, p. 425). Vitt et al. (1995) recorded it from P.N.G.
Epiphytic on a wide range of indigenous dicot trees and shrubs (at least 22 genera, and at least 3 species of southern beech) as well as Cordyline australis, and some podocarps. It is an abundant species on upper branches in montane Fuscospora forest in the eastern mountains of the South I. and probably elsewhere. It also occurs on a range of adventive tree genera, especially Populus and Salix. Usually occurring in strongly insolated sites such as forest margins, adjacent to light gaps, or in the forest canopy. On the South I. it occurs over a wide altitudinal range and equally commonly on both sides of the Main Divide. It occurs rarely on rock.
On the North I. from c. 10 m (Western Lake Road, Wellington L.D.) to at least 750 m (near Erua, Wellington L.D.); on the South I. from near sea level (Goose Bay, Marlborough L.D.) to c. 1200 m (Billies Knob, Nelson L.D., and Mt Alfred, Otago L.D.) Its most frequent associate is Calyptopogon mnioides, but the diversity of host species is reflected in a wide range of bryophyte associates, which include Leptostomum inclinans, Macromitrium spp. (including M. microstomum and M. prorepens), Orthotrichum graphiomitrium, Papillaria flavolimbata, Rhaphidorrhynchium amoenum, Syntrichia papillosa, and Tortula abruptinervis).
Rare N.Z. populations having widely spreading or squarrose dry branch leaves led to Müller’s (1898) description of Macromitrium recurvulum from "North Canterbury" using a Beckett collection. This taxon was accepted at the species level by Dixon (1926, p. 182) and at varietal level by Sainsbury (1955, p. 229). Vitt (1980) found no type material in the Beckett herbarium and designated a neotype with distinctly squarrose dry branch leaves for this name. I concur with Vitt’s observation that leaf set often varies in single plants and that M. recurvulum Müll.Hal. deserves no taxonomic recognition.
All the N.Z. material of M. tenue studied was cladautoicous; I have not observed material with perigonia occurring immediately below the perichaetium (gonioautoicous) such as reported by Vitt (1980, p. 427). Nor have I observed incurved branch leaf tips in N.Z. material such as recorded by him.