Class
Family
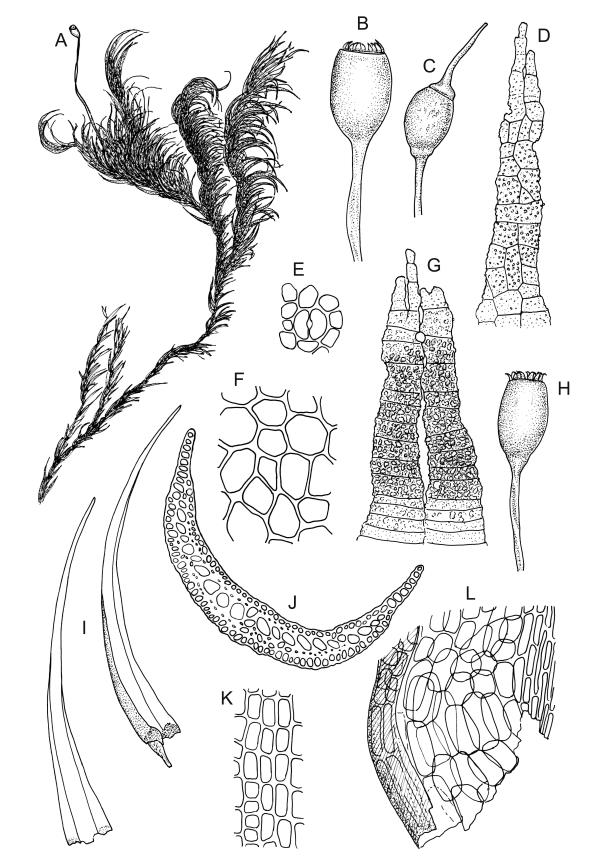
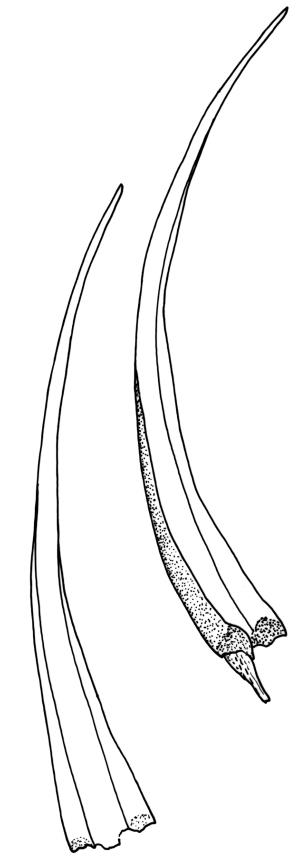
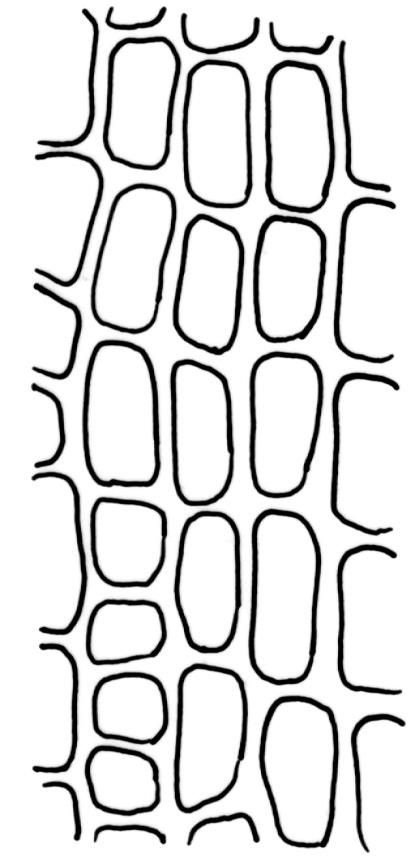
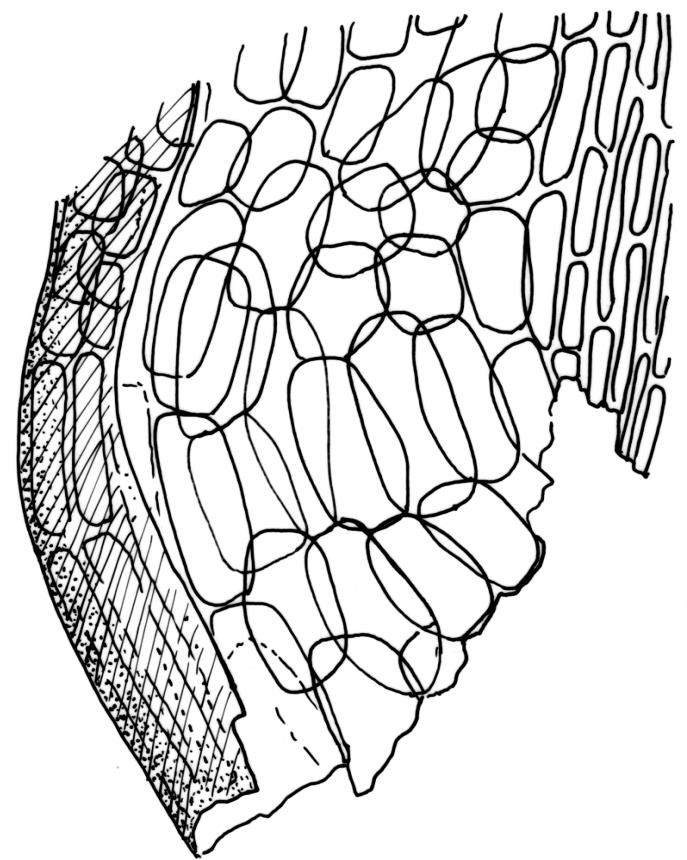
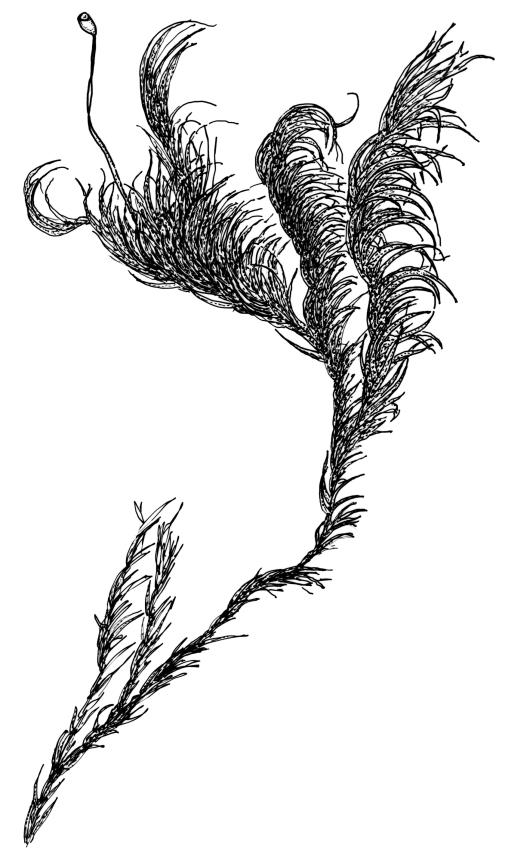
Plants robust, yellow-green when fresh and moist, becoming brown-green when dry, dull, forming extensive turves on irrigated rock. Stems c. 40–50 mm, much branched by both forking and innovation, in cross-section lacking a central strand, with 2–3 layers of small, thick-walled cortical cells, sparsely beset with red-brown, smooth, weakly branched, and ± straight rhizoids (mostly arising in leaf axils). Leaves secund and strongly curved or falcate both moist and dry, linear-lanceolate and evenly tapered throughout, ± auriculate and clasping at insertion, the distal ⅓ to ½ filled by the costa (see notes below), rounded at the apex, entire, broadly U-shaped throughout, 3.0–6.5 mm, variable in size on single stem; laminal cells (c. ⅓ above base) quadrate to short-rectangular, c. 6–12 × 5–6 µm, firm-walled, forming a broad bistratose juxtacostal band (not well illustrated in Image: Image\2RCL), unistratose in 4–5 marginal rows, with fine cuticular striations (apparently on both surfaces) that extend between cells; laminal cells closer to the leaf base becoming unistratose and more pellucid, not elongate, forming a large area of more clearly defined cells immediately above the alar cells; alar cells strongly inflated and rounded, to c. 30–40 × 25 µm, ± pigmented, forming a large, well-defined, and ± auriculate group extending to the costa and sometimes remaining attached to the stem when the leaves are removed. Costa stout, c. 290–325 µm wide, c. ⅓ to ½ the leaf width, and well-defined in lowest 1 mm of leaf, rapidly becoming ill-defined above; in surface view with short-rectangular cells on the adaxial surface and quadrate cells on the abaxial surface; in cross-section (at mid leaf) C-shaped, with a row of median guide cells and with the abaxial surface cells appearing finely papillose due to cuticular striations.
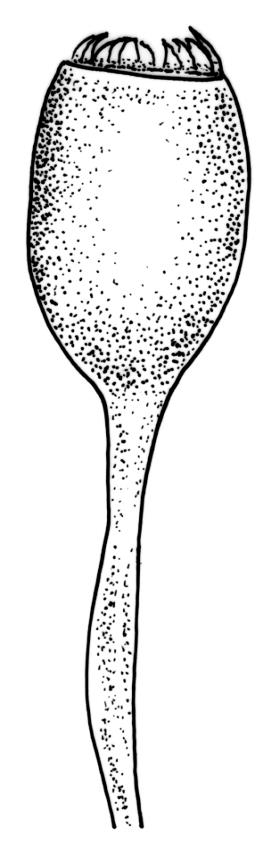
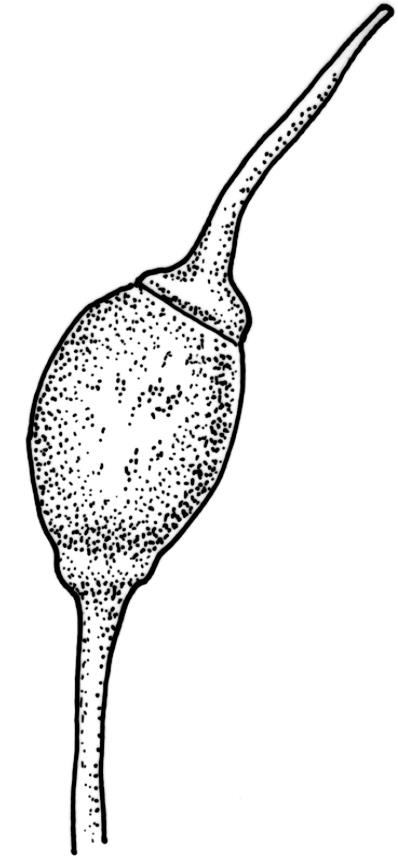
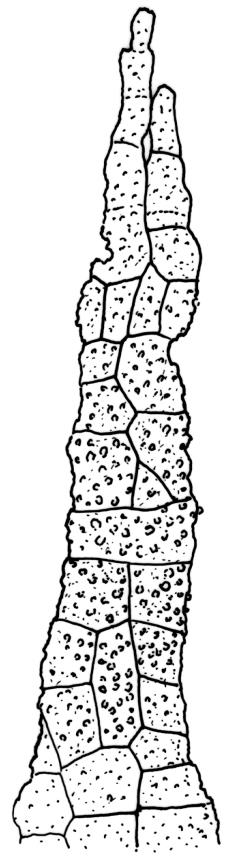
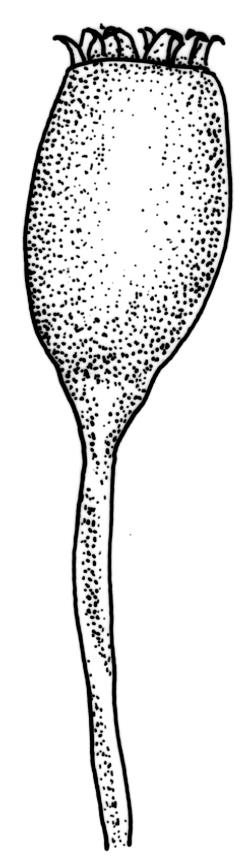
Autoicous. Perichaetia terminal but usually overtopped by innovation, often 2 or more per shoot, with perichaetial leaves differentiated, usually shorter than vegetative leaves, sheathing at base and ± distinctly shouldered. Perigonia on very short shoots at the base of the perichaetia, with the inner bracts short, oblong-ovate, and weakly costate; antheridia <10 per perigonium, mixed with filiform, uniseriate paraphyses. Setae (12–)18–23 mm, flexuose and sinistrorse, yellow-brown; capsules broadly ellipsoid and scarcely constricted below the mouth when dry, scarcely altered when moist, 1.3–1.5 × 0.8–0.9 mm, smooth; exothecial cells at mid urn rounded-polygonal, firm-walled, weakly collenchymatous, becoming isodiametric or ± oblate in several rows at the mouth; stomata restricted to capsule base, superficial; operculum long-rostrate from a conic base, oblique, ± equal the capsule in length, probably systylious. Peristome teeth well-developed, inserted close to rim, yellow-brown, very irregular in outline, often anastomosing and then the paired teeth appearing cribose, to 270 µm long × 75 µm wide (when paired, the fused teeth collectively to c. 120 µm wide), the outer surface trabeculate-lamellate, either coarsely verrucose or sometimes nearly smooth, the inner surface with an irregular median line, thicker cell walls, and moderately to strongly verrucose; preperistome absent in N.Z. material. Calyptra cucullate, smooth. Spores 22–30 µm, finely papillose, brown.
Allison 1963, figs. 1–5 (as Chorisodontium burrowsii); Ochyra 1993, figs. 1–3; Ochyra 1998, fig. 78; Cardot 1908, pl. 4 is probably also this species (cited as Verrucidens turpis).
Plants of Holodontium strictum, both fresh and dried, have a characteristic dull, yellow-green coloration and linear-lanceolate, subtubulose leaves, which are curved to falcate-secund. These features, combined with the very broad but ill-defined costa, the quadrate to short-rectangular and obscure laminal cells, and its high elevation habitat, make this a relatively distinctive, albeit very rare, plant.
Many features of H. strictum, including its irrigated habitat, are suggestive of a Blindia. However, the exserted capsule here is ellipsoid rather than the hemispheric or turbinate capsules found in the genus Blindia, where it can be either immersed or exserted. Also, the combination of the secund leaves, a broad costa, and quadrate to short-rectangular, partially bistratose laminal cells extending nearly to the insertion is not found in any species of Blindia. Finally, the presence of laminal cell wall striations and the lack of a central strand in H. strictum preclude confusion.
SI: Canterbury (Rough Creek Basin, Mt Wakefield), Otago (Old Man Range, Lake Alta on Remarkable Range).
Austral-Andean. South Georgia*, Argentina (Tierra del Fuego)*. Reported from Chile, Crozet Is, Kerguelen, South Shetland Is, and from high elevation (c. 4200 m) in Ecuador by Ochyra (1993).
Growing at alpine stream margins and at base of late snowbanks. At Lake Alta this species formed turves to c. 15 cm diam. in a flush on a steep, insolated, north-west-facing schist slope at 1900 m. A similar habitat was described (in herb.) by H.D. Wilson for a lower elevation collection from Mt Wakefield. Documented from 1520 m (Old Man Range) to 1900 m (Lake Alta).
Ochyra (1993) determined that the Kerguelen basionym of Holodontium strictum is both conspecific and earlier than the Ecuadorian type of Dicranum inerme. His conclusion that these two names are conspecific is accepted here. He provided both a detailed description and detailed illustrations of H. strictum based on type and non-type material. Although type material of these names has been seen only online for the present study, the images of this robust plant agree well with N.Z. material. Numerous British Antarctic Survey collections from South Georgia (named as H. inerme (Mitt.) Broth.) have been examined, including duplicates of South Georgian collections cited by Ochyra. Comparison to his description and illustrations permits no doubt that the ample and often fruiting N.Z. material is conspecific with H. strictum. The conspecifity of a high-alpine N.Z. species with plants occurring on both Kerguelen and the northern Andes has other examples in the N.Z. moss flora.
The placement in synonymy here of Chorisodontium burrowsii Allison (which never sat comfortably in the genus Chorisodontium) clarifies a long-standing confusion.
A likely addition to the synonymy of H. strictum is suggested by the single setae-bearing specimen of Verrucidens turpis (Cardot) Cardot (from Ushuaia, Argentina, and named by C. Matteri) in CHR. The basionym of this name, Blindia turpis Cardot, was described from Tierra del Fuego and subsequently transferred to the monotypic genus Verrucidens by Cardot (1908, p. 85). Type material of this species name has not been available for study. However, the Ushuaia specimen so-named (Musci Fuegiana Exsiccati no. 15, CHR 457017!) compares extremely well to published illustrations and to N.Z. and South Georgian material. All have falcate-secund leaves, subquadrate laminal cells, rounded leaf apices, and strongly differentiated, inflated alar cells. The area of juxtacostal bistratose cells is less well-developed in the Patagonian material. The likelihood that Verrucidens turpis (Cardot) Cardot is conspecific with H. strictum deserves further investigation.
The nature and extent of the costae in H. strictum are difficult to interpret. The costae in the basal c. 1 mm of the leaves are relatively clearly defined and c. 290–325 µm wide, with the adjacent laminal cells pellucid and unistratose. Approximately 1 mm above the insertion, the definition of the costae becomes more obscure. This is partly because of the U-shaped leaf cross-section, but is also due to the narrow and partially bistratose lamina (c. 10–15 cells on each side of the costa, inadequately illustrated here), becoming increasingly opaque. At mid leaf the lamina is roughly 6–8 cells wide, including the bistratose portion, and the boundary between the costa and the lamina is indistinct, even in cross-section. When viewed under the stereoscope the lamina appears to extend ½ to ⅔ the total leaf length. Ochyra (1993, figs 16–24) provided an excellent and detailed series of costal illustrations.
The laminal cells (c. ⅓ above base) bear fine cuticular striations that extend across cell boundaries; these striations can be difficult to interpret under the light microscope but they are similar to those found in some species of Amphidium. They can be readily observed in leaf cross-sections.
The ornamentation of the peristome teeth is also very difficult to interpret with the light microscope and seems to vary between populations. The verrucae are interpreted here as occurring on both surfaces of the teeth. The outer surface of the tooth has thinner walls (best seen in cross-section) and bears transverse lamellae. The development of the verrucae on the outer surface appears to vary between populations.