Class
Family
Genus
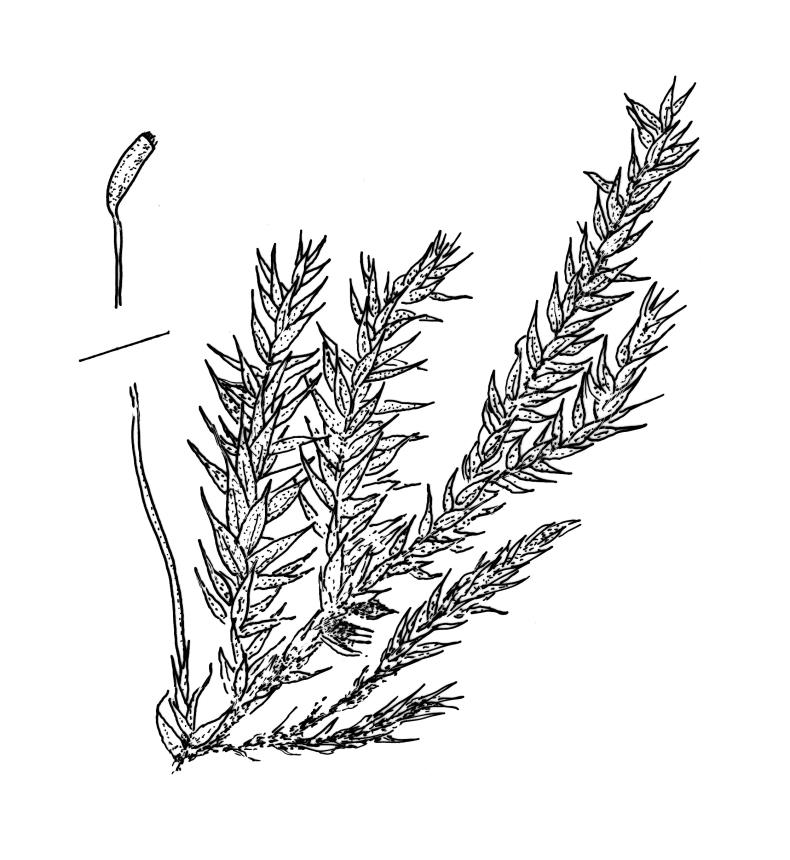
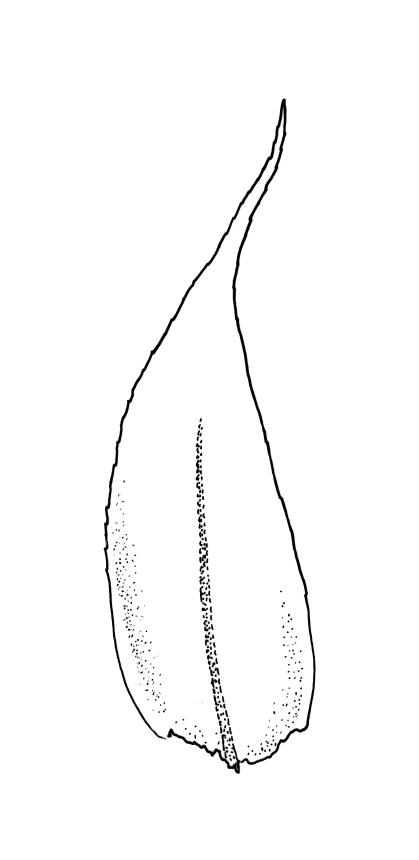
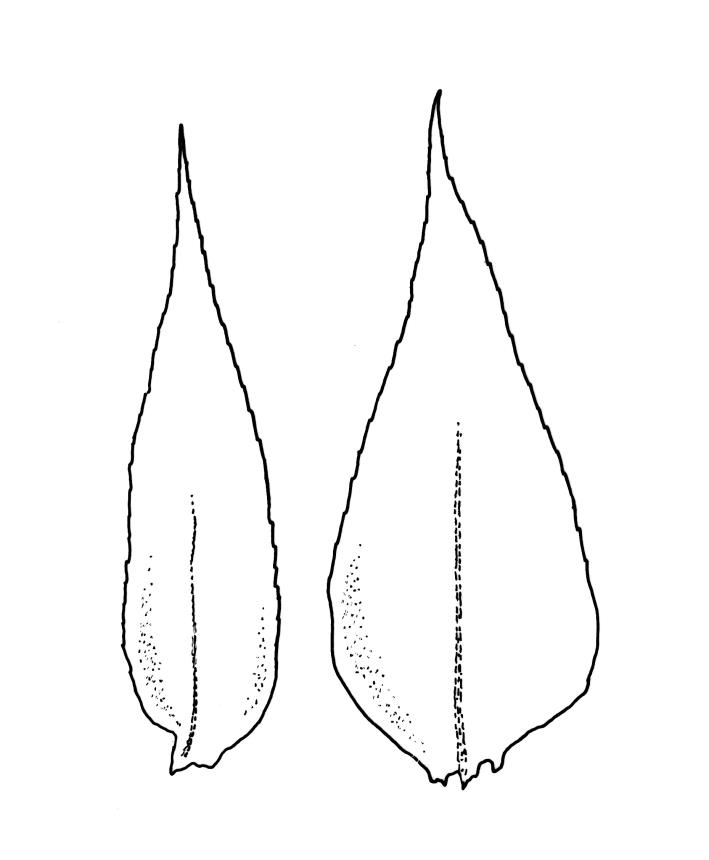
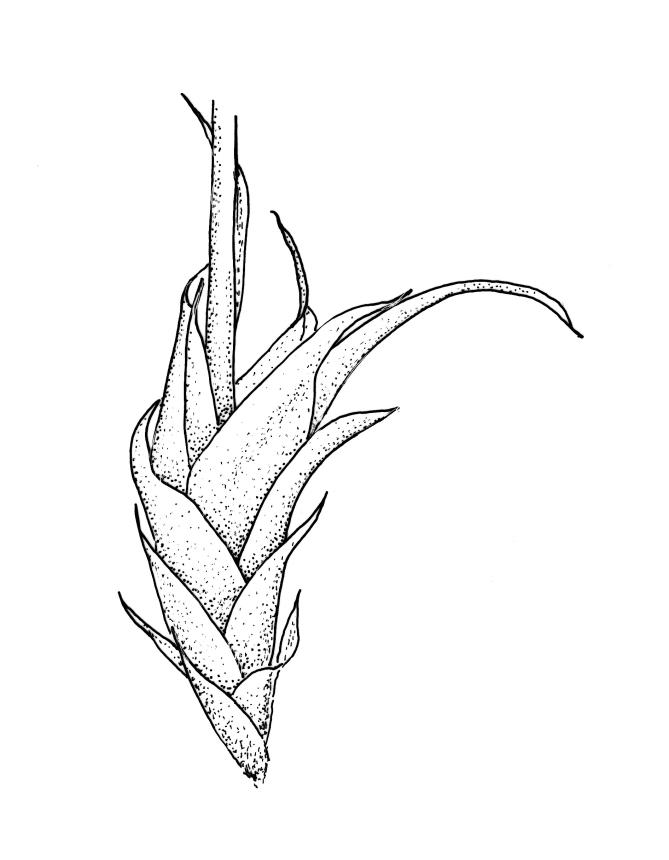
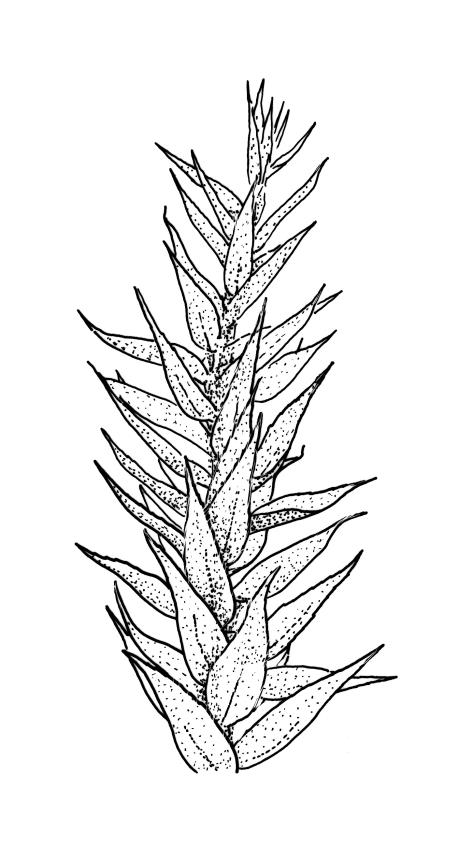
Plants yellow- or bright green, lustrous, forming interwoven mats on bark. Stems irregularly branched, to at least 30 mm long, in cross-section with 3–5 layers of thick-walled outer cells and a distinct central strand; rhizoids smooth, red-brown, scattered in fascicles on ventral surface of the stem. Branches variable in length and often forked, 3–12 mm. Stem and branch leaves differing only by size, little altered when dry. Stem leaves spreading complanate, ovate-lanceolate and nearly evenly tapered to an acuminate apex, weakly concave, narrowed to the insertion, ± symmetric, plane and denticulate to base, neither decurrent nor striolate, 1.4–1.45 × 0.45–0.6 mm. Branch leaves slightly smaller, mostly 1.2–1.4(–1.6) × 0.3–0.5 mm, not otherwise differentiated. Costa weak and tapered, mostly extending c. ½ or slightly greater leaf length, lacking an abaxial terminal spine. Upper laminal cells linear, smooth or slightly prorate, mostly 57–65(–75) × 4–6 µm; cells of the acumen slightly shorter; basal cells shorter and thicker-walled; alar cells irregular to ± subquadrate, in a small and poorly differentiated group.
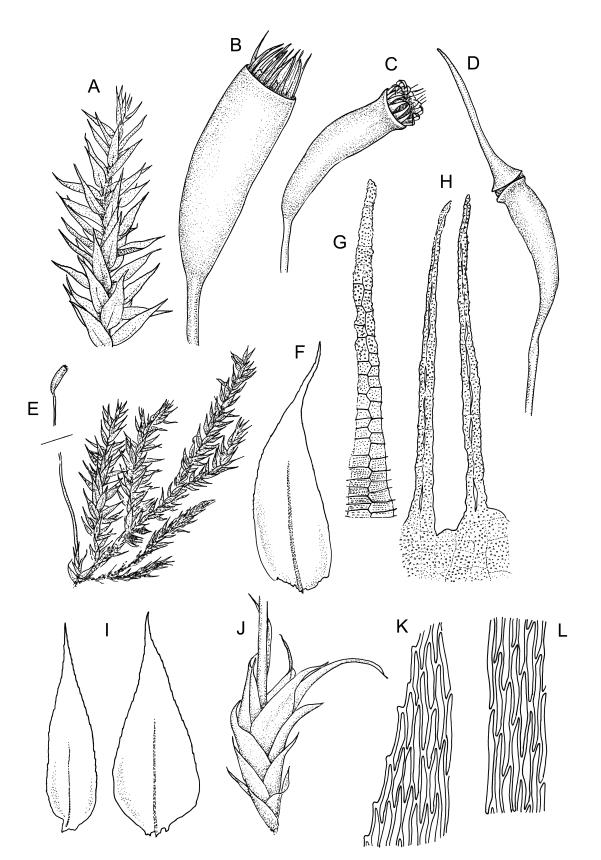
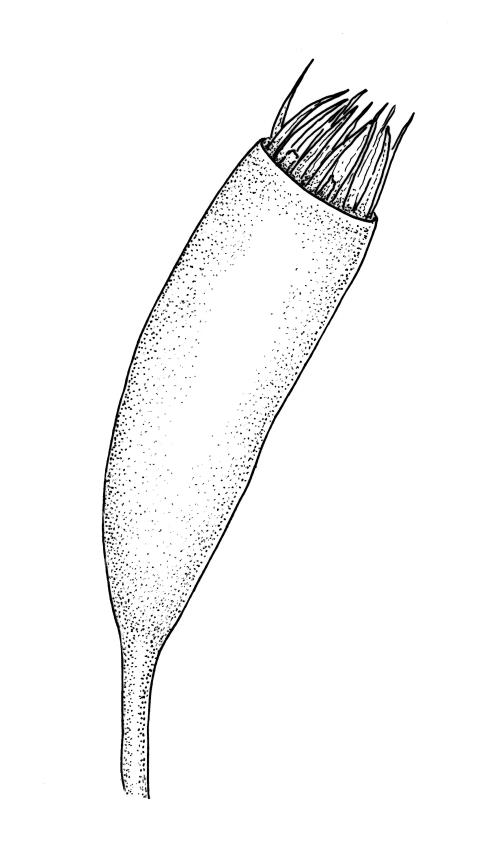
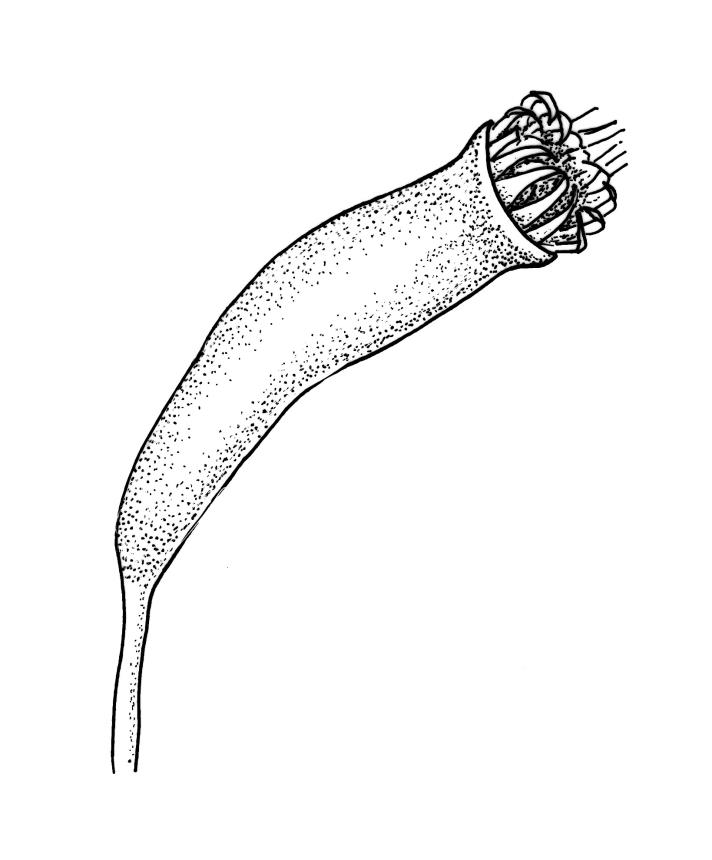
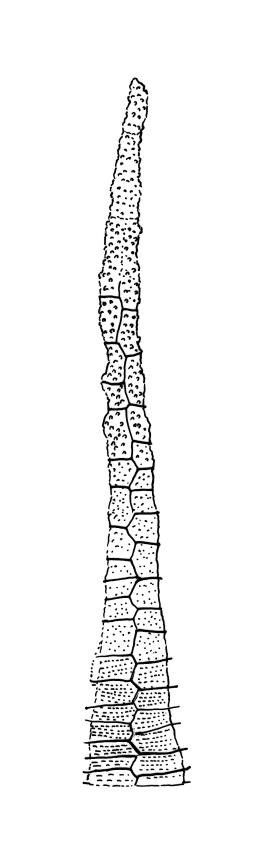
Autoicous. Perichaetia scattered on main stem, c. 2–2.5 mm long, sheathing the lower seta, the leaves abruptly acuminate from an oblong base, erect, ecostate. Perigonia scattered and usually numerous on fruiting stems, c. 0.8 mm, with filiform paraphyses. Setae slender, not twisted, mostly 20–25(–30) mm, red-brown, smooth; capsules narrowly cylindric, variable in length, curved, red-brown at maturity (but often green at dehiscence), narrowed below the mouth when dry, inclined to horizontal, 1.6–2.6 mm; exothecial cells oblong, firm-walled, not collenchymatous; stomata not seen; annulus weakly differentiated; operculum long rostrate from a conic base, curved, c. 1.2–1.6 mm and c. 0.6–1 × the length of the theca. Exostome teeth pale yellow, usually reflexed at base and circinate when dry, linear, bordered but not shouldered, coarsely papillose-baculate throughout on the outer surface, the papillae mostly transversely aligned on the lowermost plates, weakly trabeculate on inner surface, c. 500–550 µm; endostome from a low (c. 120 µm) membrane, with segments linear, nearly the height of the teeth, and cilia either absent or poorly developed. Calyptra cucullate and smooth. Spores 15–21 µm, green, rather coarsely insulate.
Sainsbury 1955, pl. 73, fig. 1; Buck 1981, figs 8–13.
Regionally, fertile material of E. cylindritheca, because of the distinctive narrowly cylindric capsule, very long and slender operculum, and distinctive peristome, is unlikely to be confused with other species. However, sterile material, which is fortunately uncommon, could easily be confused with Rhynchostegium muriculatum. Eriodon cylindritheca has branches that are decidedly more complanate, and stem leaf mid to upper laminal cells generally shorter (57–75 µm vs mostly 75–90 µm) than those of R. muriculatum.
K; NI: N Auckland (Waipoua), S Auckland, Gisborne, Hawke’s Bay (Dannevirke), Taranaki (near Dawson Falls, Pōtaemā Swamp Track), Wellington; SI: Nelson, Westland, Otago (Mossy Falls, Chaslands), Southland; St.
Australasian. Reported from mainland Australia (N.S.W.) by both Dixon (1929) and Hedenäs (2002), apparently on the basis of a single collection.
Epiphytic and usually on twigs, small branches or trunks of small diameter; also occurring on fallen logs and usually close to streams. Rarely on shaded rock. Occurring on a wide range of host species, including Aristotelia serrata, Coprosma grandifolia, C. rotundifolia, Fuchsia excorticata, Hoheria glabrata, H. sexstylosa, Melicytus ramiflorus, Metrosideros diffusa, Muehlenbeckia australis, Myrsine divaricata, Ripogonum scandens, Salix sp., Sambucus nigra, Schefflera digitata, Urtica ferox, and Weinmannia sylvicola. Occurring with Calyptopogon mnioides, Cryphaea tenella, Neckera pennata, Rhaphidorrhynchium amoenum, Sauloma tenella, and Weymouthia cochlearifolia. The presence of the relatively rare Daltonia splachnoides in many herbarium collections is noteworthy. Occurring from near sea level (Fox River, Nelson L.D.) to at least 480 m (Lake Rotoroa, Nelson L.D.) on the South I. and to c. 1070 m (Mt Manuoha, Gisborne L.D.) on the North I. On the South I. this species appears to be absent from Marlborough and Canterbury L.D. as well as drier parts of Otago L.D. On the North I. the only confirmed Hawke’s Bay material is an 1887 Colenso collection from Dannevirke.
More precise collection data than recorded on WELT M016047 (an isosyntype) of Dixon’s basionym are given in the protologue. The protologue records the date of collection as 26 Sept. 1912 and cites what appears to be a W. Gray collection number (145). WELT M016047 has a Sainsbury herbarium number (9398) and was sent to Sainsbury by Dixon. Apart from one poorly documented and sterile isosyntype in CHR, none of the other syntypes cited by Dixon appear to be represented in N.Z. herbaria. For this reason, a duplicate of W. Gray 145, if present in the Dixon herbarium, would be the preferred choice as a lectotype. Hedenäs (2002) cited an apparent duplicate of W. Gray 145 in H‑Brotherus as a syntype.
Hedenäs (2002) placed the Australian Helicodontiadelphus australiensis Dixon [J. Bot. 74: 5, 1936] into synonymy here, based on type material collected by R. Collie in N.S.W. Material collected by Collie was also cited by Dixon (1929, p. 329) in his discussion of this species.
There is some confusion about the species relationships and consequent nomenclature in Eriodon. Based on examination of a single ample specimen of the generitype, E. conostomus (Dusén 372, CHR 585851), I agree with Buck’s (1981) suggestion that E. conostomus and E. cylindritheca are not conspecific.
Buck (1981) referred the Andean E. radicalis Spruce ex A.Jaeger to the genus Entodontopsis. However, an isotype from Ecuador (Musci Amazonici et Andini no. 1401; CHR 414932) is clearly an Eriodon, and if this material were collected in N.Z. it would be almost certainly referred to E. cylindritheca. The peristome teeth are c. 600 µm long and the spores mostly 16–20 µm and coarsely ornamented. The alar cells in the Ecuadorian collection are scarcely differentiated rather than forming a large group of subquadrate cells as illustrated by Buck (1981, fig. 15). The species-level taxonomy of Eriodon requires further study beyond the scope of this eFlora.