Class
Family
Genus
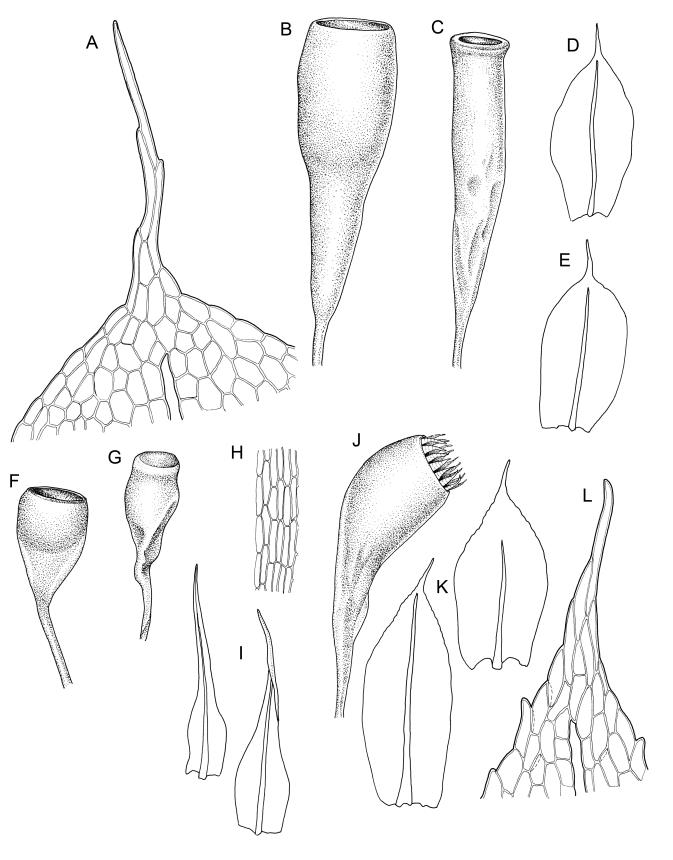
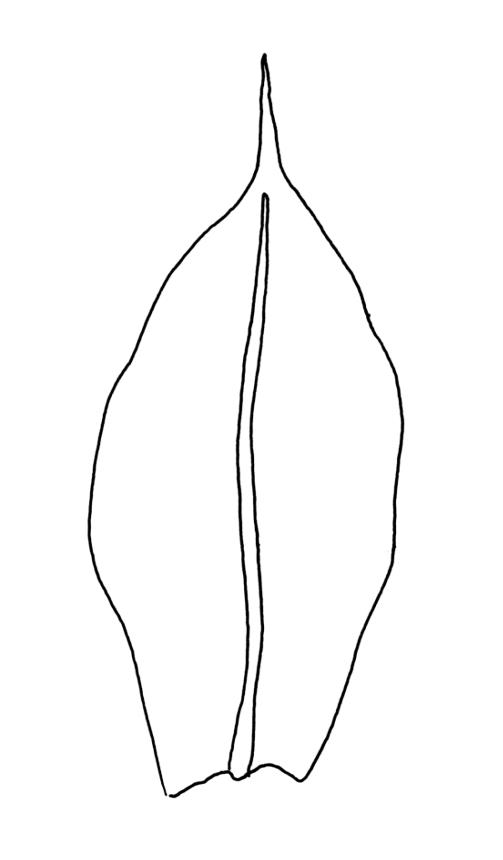
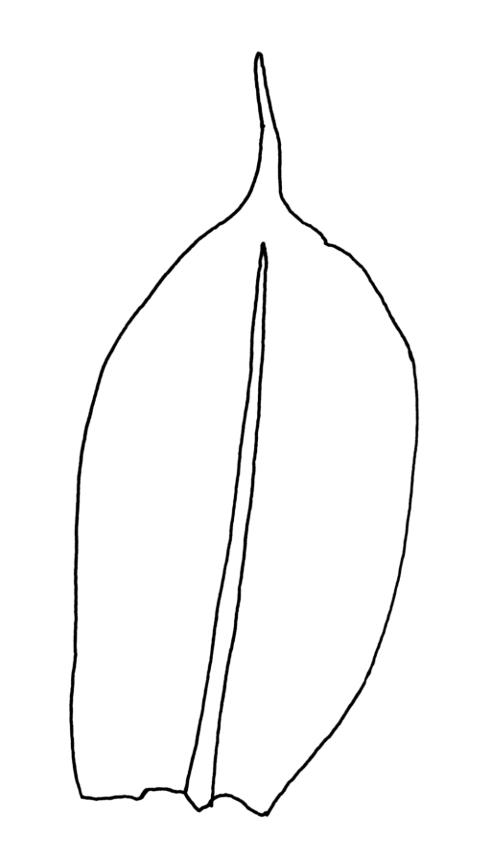
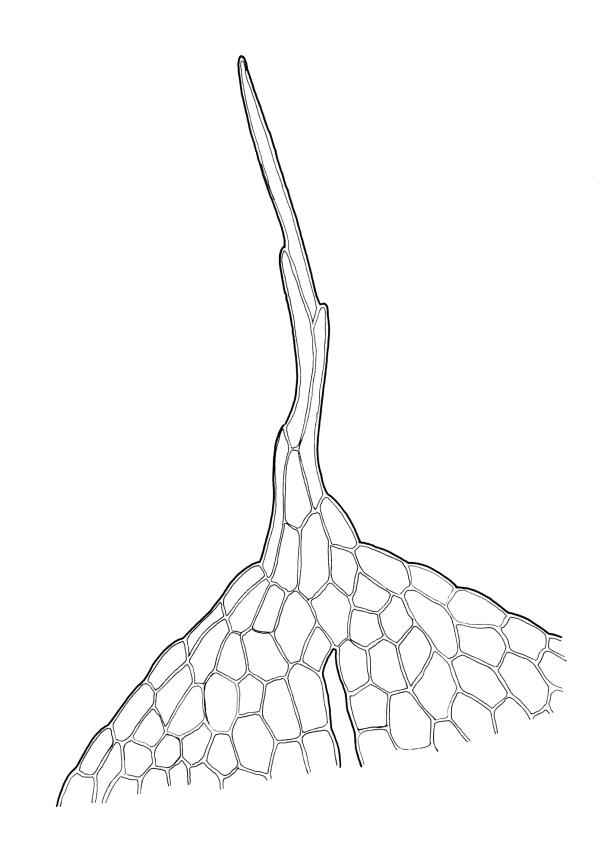
Plants yellow-green. Stems red-brown, 2–5(–7) mm high, branching once by subperigonial innovation, beset with red-brown rhizoids. Leaves erect-spreading, broadly obovate to oblong-spathulate, 1.5–3.5 × 0.75–0.9 mm (including arista), concave, entire, tapered in upper ⅓ to a slender, ± yellow arista or very rarely with arista absent (see below); upper laminal cells oblong-hexagonal, (24–)30–60 × 17–30 µm, longer and more oblong below; marginal cells not differentiated; apical cell (150–)180–300(–400) µm; alar cells 2–4, moderately inflated. Costa c. 30–50 µm wide near base, failing a few to several cells below base of the arista or fusing with it and appearing long excurrent. Axillary hairs present.
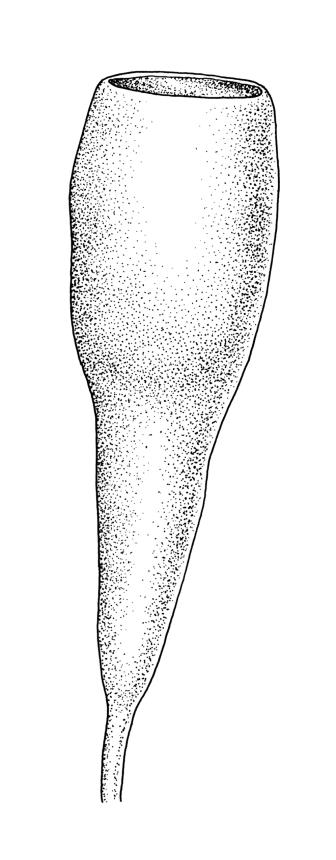
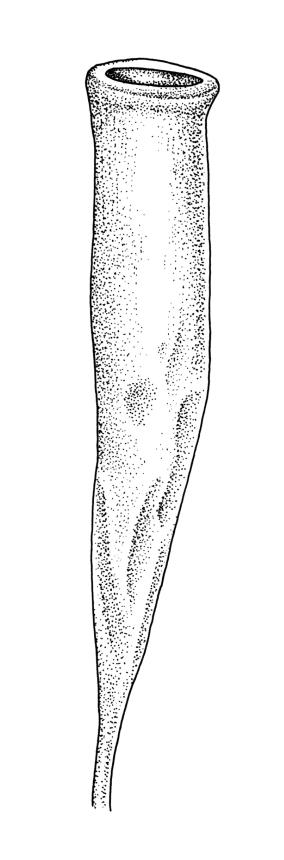
Autoicous. Perigonia and perichaetia as per genus. Setae 2–4(rarely–7) mm, weakly dextrorse, weakly hygroscopic, pale red-brown; capsules erect or rarely inclined, symmetric, oblong-cylindric, constricted below mouth when dry, (2.0–)3.0–4.5 mm, with a well differentiated, tapered neck c. ½ the capsule length, red-brown; mouth ⅔ to equal the capsule diameter, transverse; exothecial cells c. 5–8:1, 50–90 µm long, with thick, strongly cuneate cell walls, c. 6–8 rows quadrate to oblate and strongly pigmented at mouth; operculum plano-convex. Peristome absent or very rarely double and weakly developed (see below). Spores (27–)30–43 µm, nearly smooth or sometimes insulate, often with trilete scars or persisting in tetrads, often collapsed when dry.
Brotherus 1924, fig. 274 (as Funaria apophysata); Catcheside 1980, fig. 122 (as Funaria apophysata); Meagher & Fuhrer 2003, p. 151.
In its most representative form, E. apophysatus has leaves similar in shape to those of E. muhlenbergii, but that species has toothed leaf margins, more markedly differentiated marginal cells, and asymmetric, peristomate capsules. Entosthodon apophysatus differs from the Australian E. subnudus var. subnudus in leaf form and in apical cell, seta, and capsule lengths, peristome development, and spore ornamentation.
NI: N Auckland (near Cascades, Erin Point, Remuera, Wiri, near Mangōnui,), S Auckland (near Huntly (aberrant)), Hawke’s Bay (Te Awanga, Havelock North, near Waipukurau (aberrant), Napier); SI: Nelson (Pūponga), Canterbury (Port Hills, Quail I.), Otago (Glendhu Bluff at Lake Wānaka).
Austral. Tasmania*, Australia (all states)*, Uruguay*, Chile*.
On damp mineral silt or clay soil, in natural or highly modified grassland or scrub; sometimes in horticultural sites (e.g. vineyards). Frequent associates include Ceratodon purpureus, E. radians, E. subnudus var. gracilis, Physcomitrium pyriforme, Pleuridium nervosum, Schizymenium bryoides, Tortula truncata, Weissia austro-crispa, and W. controversa. Ranging in elevation from sea level (Wiri) to c. 300 m (Lake Wānaka).
The long, oblong-cylindric, gymnostomous capsules, short setae, and markedly aristate comal leaves in most instances set this species apart from all other N.Z. species in the genus. However, E. apophysatus in N.Z. is variable in both sporophyte and gametophyte features and a small number of aberrant collections have been seen. The sporophytic variability here is consistent with high levels of plasticity in other Entosthodon species and is arguably consistent with the high levels of variability and evolutionary homoplasy hypothesised for the genus as a whole by Liu et al. (2012).
In aberrant material from near Huntly, S Auckland L.D. (J.K. Bartlett 5169; CHR 413349), some sporophytes are gymnostomous while others are peristomate. The gymnostomous plants have setae to 5 mm (slightly longer than usual for E. apophysatus), aristate leaves, ± tetrahedral spores, and are thus representative of the species. However, other plants in the same collection have weakly developed double peristomes, longer than usual setae (to 7 mm), comal leaves mostly acute or with a very short aristae, and costae ending below the base of the aristae. The peristome teeth here are highly irregular, striate, mostly broadly rounded apically, and c. 75–150 µm high, and the endostome segments are irregular and to c. 90 µm high. Spores throughout the collection appear to be normally developed for E. apophysatus, mostly 27–36 µm, weakly tetahedral (with three ± flat surfaces), apparently insulate, occasionally trilete and with a few persisting as tetrads.
Aberrant material from near Waipukurau, Hawke’s Bay L.D. (R.E. Beever s.n., 19 Oct. 2009, CHR 604767) has consistently non-cuspidate comal leaves but representative and entirely gymnostomous capsules. The unusual form of the leaves here may be a result of herbicide spraying in a vineyard.
A single collection from eastern Spirits Bay, N Auckland L.D. (J.K. Bartlett 22431, CHR 91832), at the northern extremity of the North I., tentatively referred to E. apophysatus, has setae much longer than typical (9–11 mm), and may deserve varietal status. However, the obovate-aristate, entire leaves, the nearly smooth, trilete spores, and long oblong-cylindric capsule indicate its affinity to this species. A very rudimentary (or possibly fragmentary) peristome has been noted in one capsule of this collection. No other material of Australasian E. apophysatus has been seen with such elongate setae.
At this stage in our understanding of E. apophysatus there seems no alternative but to record aberrant material and to acknowledge the puzzling variability of this taxon.
Entosthodon clavaeformis Müll.Hal. & Hampe (South Australia), Funaria aristata Broth. (N.S.W.), and E. clavellatus Mitt. (Uruguay) are synonymous with E. apophysatus.