Class
Family
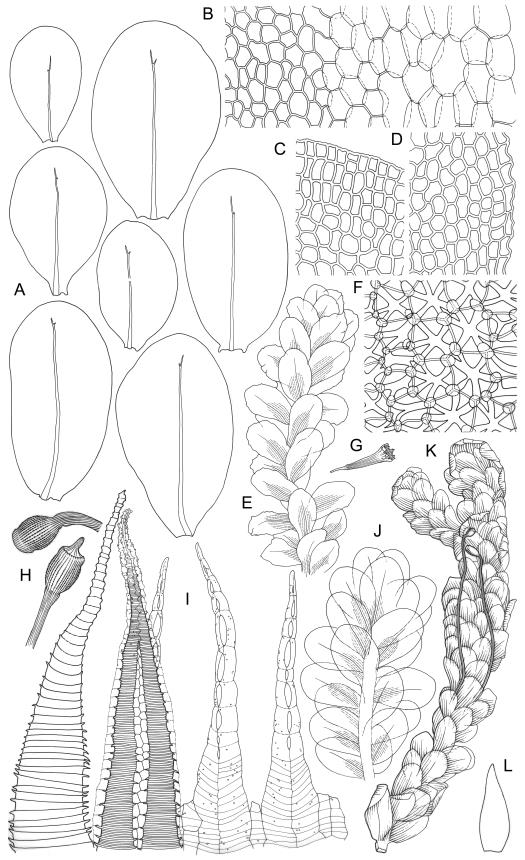
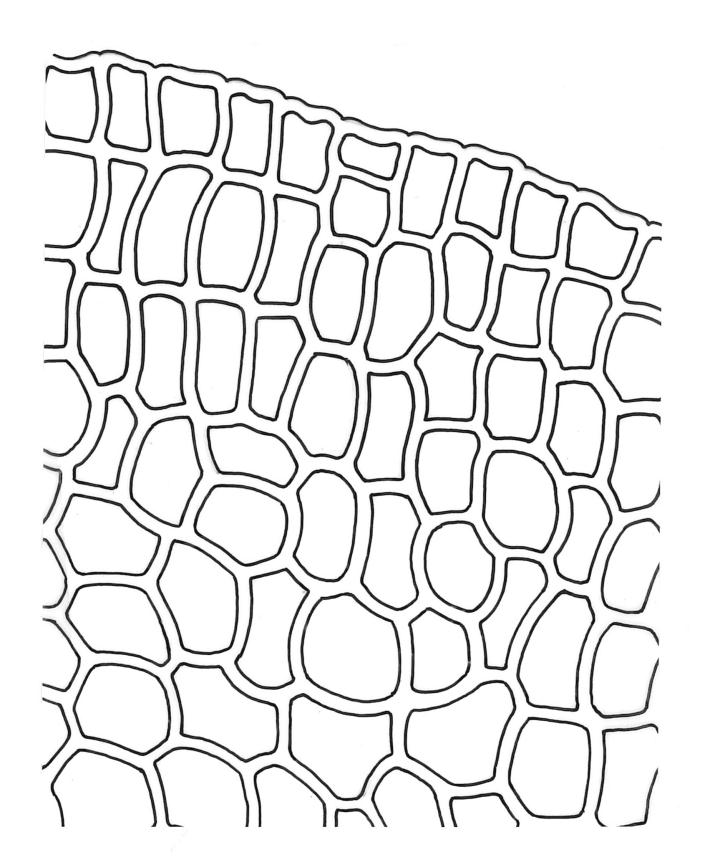
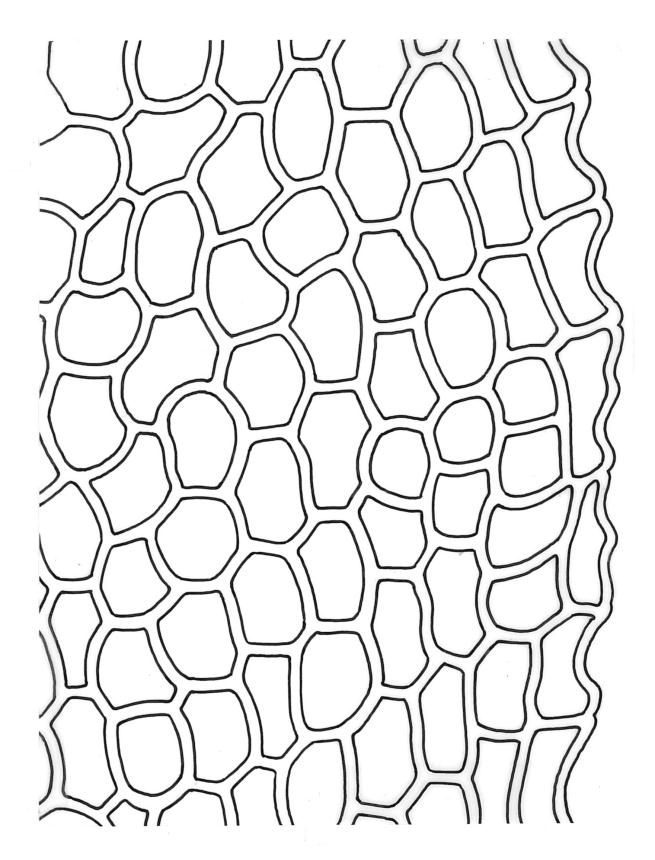
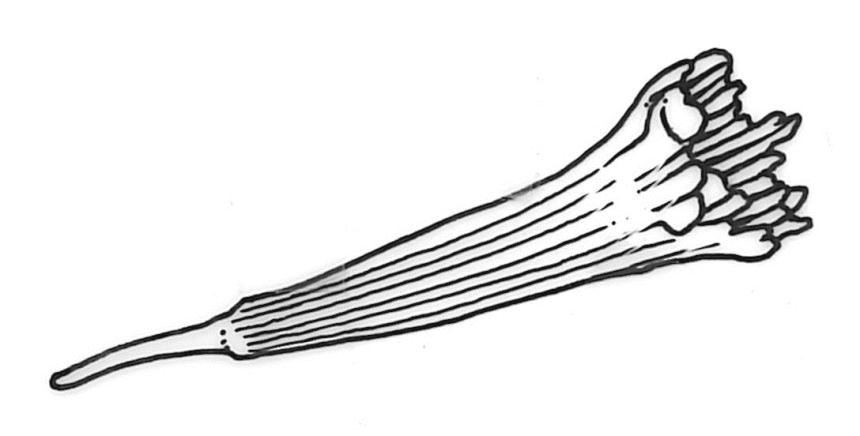
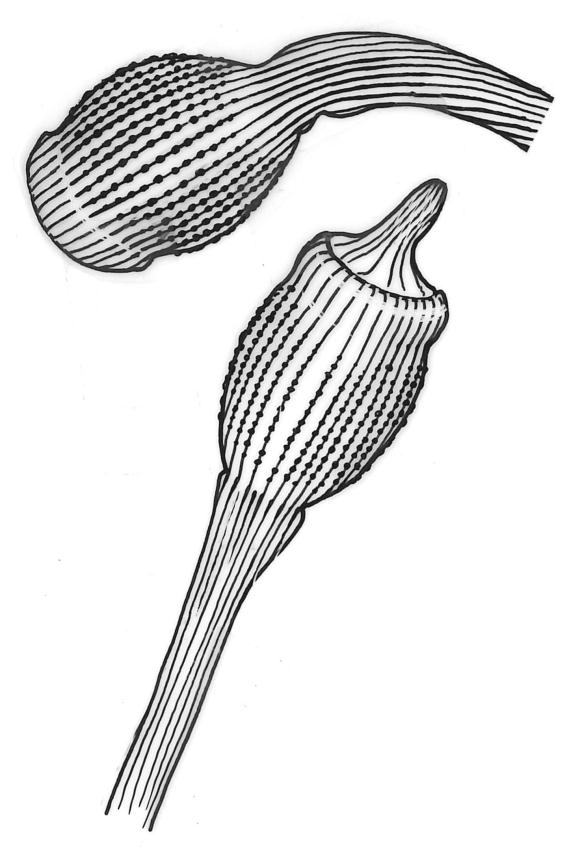
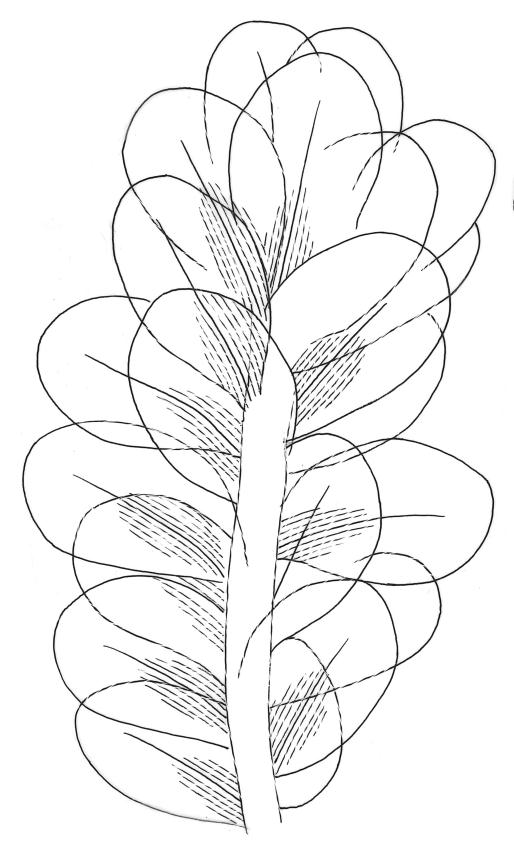
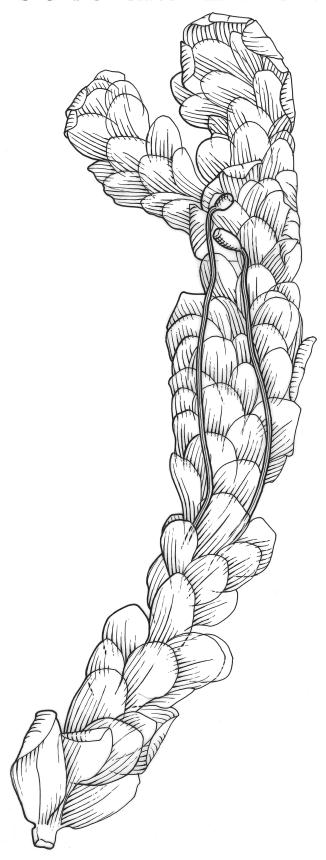
Plants robust or medium-sized, yellow-green, often tinged with brown (never red) pigments, scarcely altered and not or weakly iridescent when dry, forming rather loose, distinctly layered mats. Stems moderately and irregularly branched, brown, to c. 80 mm (rarely as short as 3 mm) long, in cross-section lacking a hyalodermis and central strand, with 3–4 layers of thick-walled, pigmented and ± well-defined cortical cells, beset below with brown, smooth rhizoids. Shoots mostly 3–4 mm wide. Leaves inserted in c. 6 ranks, complanate, somewhat asymmetric (especially at insertion of lateral leaves), not or little crisped when dry, obovate to oblong-elliptic from a narrowed insertion, broadly rounded at apex, not apiculate, plane, unbordered, finely crenulate throughout, (1.0–)2.5–4.0 × (0.8–)1.1–2.5 mm; upper laminal cells firm-walled, irregular to ± hexagonal, 12–18(–21) × (6–)8–10(–15) µm; juxtacostal cells greatly enlarged (to c. 120 × 36 µm), lax, ± hexagonal, forming a large but ill-defined group extending c. ⅔ the length of the costa and ⅓ to ½ to the margin (clearly visible with a hand-lens); marginal cells not elongate, somewhat smaller and more quadrate than adjacent laminal cells but not forming a distinct border. Costa yellow-brown, c. 30–75 µm wide in lowest ⅓, unbranched, spurred, or branched near terminus, extending ¾ or more the leaf length and usually ending 200–350 µm below leaf apex.
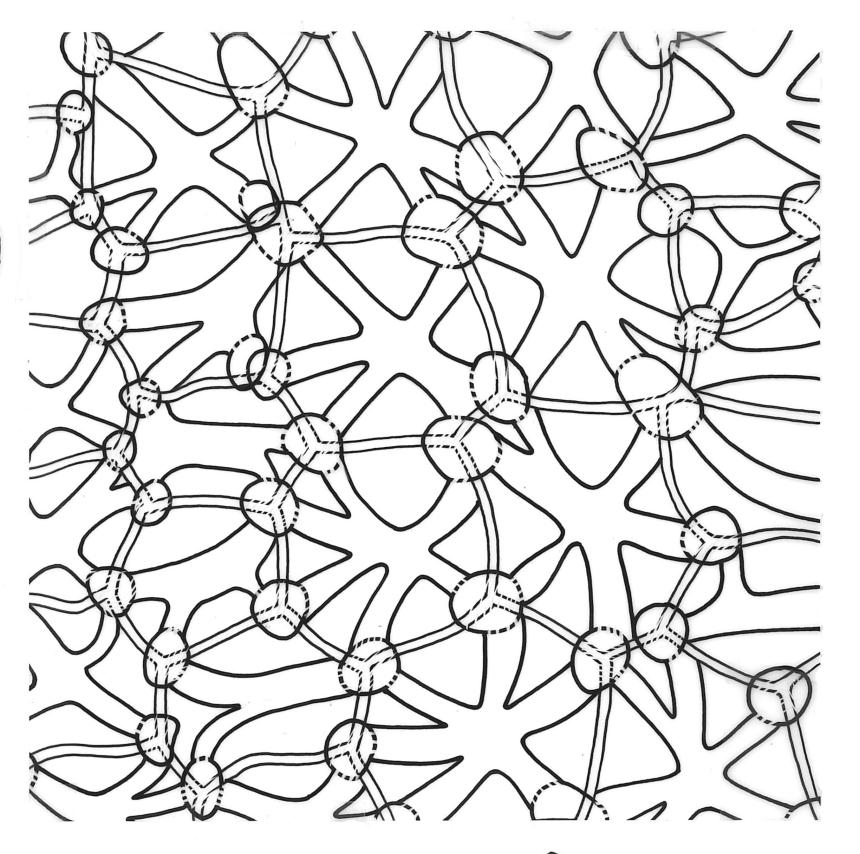
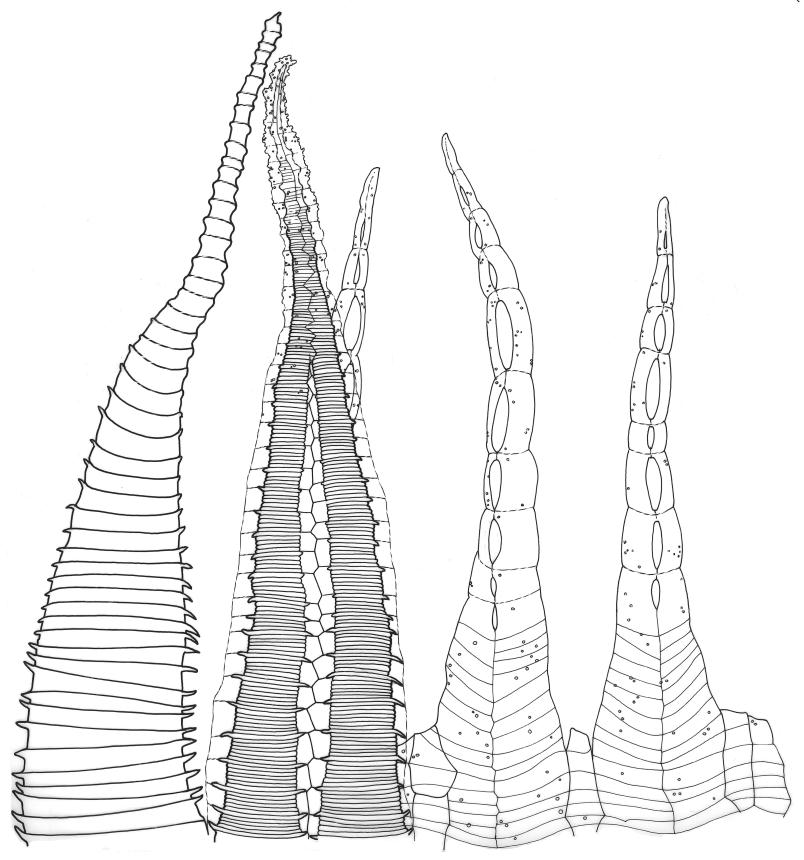
Dioicous. Perichaetia scattered on ventral surface of stems, with perichaetial leaves ovate, acute, ecostate, and c. 800 µm long. Perigonia gemmiform, scattered on ♂ stems, with ovate, acute, ecostate bracts c. 650 µm long, enclosing c. 4–5 antheridia, and lacking paraphyses. Setae arising from ventral surface of stem and inconspicuous, 5–9(–13) mm, smooth, weakly sinistrorse, red-brown; capsules inconspicuous (mostly hidden by shoots in fresh material), erect to horizontal, ovoid or obovoid, c. 1 mm long, with a weakly developed neck, not constricted below the mouth when dry, appearing scabrous under hand-lens, dark red-brown; exothecial cells ± hexagonal, 30–56 µm in greater diam., with collenchymatous secondary thickenings and (5–)6(–11) columnar arching secondary thickenings that arise from the cell angles and converge on a central point over the lumen to form a ribbed vault-like structure; annulus ill-defined; operculum rostrate from a conic base, c. 0.7 mm long. Exostome teeth c. 360 µm long; endostome segments arising from a moderately high basal membrane, equal to the teeth in height (c. 165 µm long excluding the membrane), nearly smooth, narrowly perforate. Calyptra mitrate-rostrate, c. 1.0 mm long, smooth at apex. Spores 9–12 µm, green, smooth.
Beever et al. 1992, fig. 68 h–i; Malcolm & Malcolm 2006, pp. 184, 238, 266.
K; NI: North Auckland, including offshore islands (LB, GB, RT), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington including KA; SI: Nelson, Marlborough, Canterbury, Westland (Lake Kaniere, near Fox Glacier, near Franz Josef, Te Naihi River), Otago, Southland; St; Ch; A. Reported from C by Vitt (1974).
Australasian. Tasmania* and mainland Australia (N.S.W.*). Recorded from many localities in Vic. by Streimann (1999) and from eastern parts of S.A. by both Catcheside (1980) and Streimann (1999).
Throughout the main islands, from lowland to upper subalpine zones. On the North I. ranging to at least 670 m (Oroua Valley, Wellington L.D.) and on the South I. ranging from 20 m or less (Ship Cove and Napenape Scenic Reserves, both Marlborough L.D.) to c. 1200 m (Mt Owen, Nelson L.D.). Common substrates include soil banks and rock outcrops, usually those associated with seepages. Also found on rotten logs, and occasionally on rocks in stream beds, and on damp forest litter. It can occur at the mouths of limestone caves or at the bases of dripping limestone outcrops and there often becomes heavily encrusted with lime. It is more common in eastern regions of both main islands than other species of Distichophyllum and appears to be an uncommon species in Westland L.D.. Its widespread occurrence in drier and more easterly parts of the South I. (and probably the North I.) is paralleled by its widespread occurrence in southern Australia, where its range extends westward to South Australia.
Although D. microcarpum is highly variable in stature, its inflated group of juxtacostal cells, normally readily seen with a hand-lens, and lack of a leaf border are constant features. The leaf areolation is highly characteristic in Distichophyllum. The juxtacostal group often becomes more conspicuous in dried material. When fresh the plants have a distinctive dull and oily appearance and yellow-brown coloration quite unlike any other N.Z. species of Distichophyllum. The usually large size of the plants, the pale appearance of the juxtacostal cell region, lack of elongate marginal cells, the placement of the perichaetia on the ventral surface of the stems, and the usual occurrence of this species in seepages make it easily recognisable in the field. In fresh material the sporophytes are inconspicuous and normally not visible until the plants are inverted. The ‘ribbed vault’ columnar secondary thickenings of the exothecial cells are another striking feature of the species. These ribbed vault thickenings cause the exothecial cells to be strongly mammillose and hence the dry capsule appears roughened under low magnification. I am unaware of similar exothecial cells elsewhere in the Daltoniaceae or related families. Collectively these features could justify the creation of a monotypic genus to accommodate D. microcarpum, but this departure from traditional placement is not advocated here. The distinctiveness of D. microcarpum within Distichophyllum was recognised by Brotherus (1925) and may be appreciated by perusal of his generic treatment, where it is placed in a small group of dioicous species within the section Mniadelphus; all other N.Z. taxa are placed by him in section Discophyllum.
The selection of a lectotype for Hypnum microcarpon Hedw., Sp. Musc. Frond. 244 (1801) is problematic. There is strong reason to believe that the "Insulae Australes" referred to in Hedwig’s protologue is N.Z. (Fife 1996). However, there appears to be neither J. Banks nor A. Menzies (the only significant collectors of N.Z. mosses prior to 1801) collections in either BM or the Hedwig-Schwägrichen herbarium at G (Fife 1996). Both the protologue and illustrations in Hedwig (1801, Tab. LIX, 6–8) are uninformative and my earlier suggestion (Fife 1996, p. 46) that typification using Hedwig’s illustration would be appropriate is therefore not acted upon here. Hopefully an appropriate lectotype for this Australasian species can be found.
As comments by Brotherus (1925, p. 231) suggest, it is unlikely that any of the species placed by him near to D. microcarpum deserve recognition. Three names based on New Zealand types (two described by Colenso and one by Mueller) are considered by Brotherus "kaum spezifisch verschieden"; type material of two have not been located. Distichophyllum levieri (Geh.) Broth. is based on material from Mt Wellington, Tasmania. Four specimens so-named by Brotherus, including one from Mt Wellington, have been studied and all are clearly referable to D. microcarpum; the type specimen has not been located. Hookeria subrotunda Hampe collected from Mt Disappointment (Victoria?) by F. Mueller (Type: NY-Jaeger!), is synonymous. I have seen no material of D. complanatum (Hampe) Mitt., which is based on a Victorian type. Distichophyllum aloma Müll.Hal. is mentioned under "excluded taxa", above.
Sainsbury’s (1949) description of Distichophyllum microcarpum var. homodictyon emphasises supposedly larger upper laminal cells and less differentiated juxtacostal cells. The upper laminal cells in the type specimen range from 15–21 × 9–15 µm, and fall at the upper end of the range of continuous variation of the species. The degree of differentiation of the juxtacostal cells likewise is not remarkable in the species context. The var. homodictyon does not deserve taxonomic recognition.