Class
Family
Genus
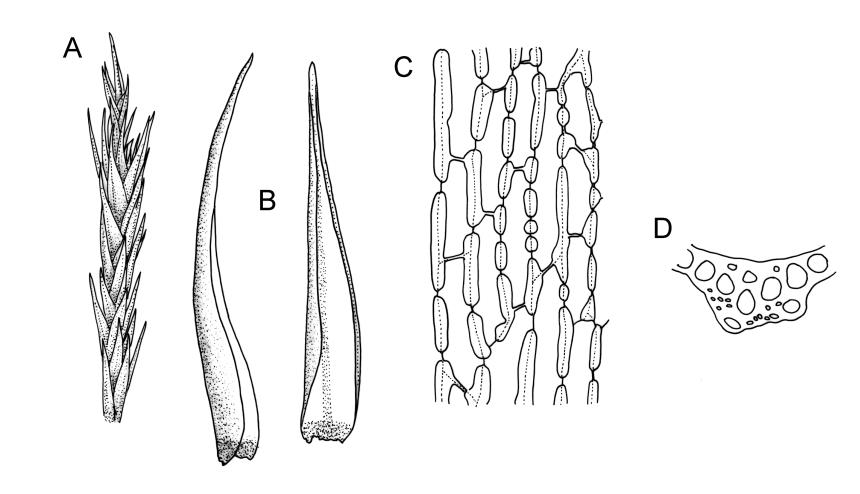
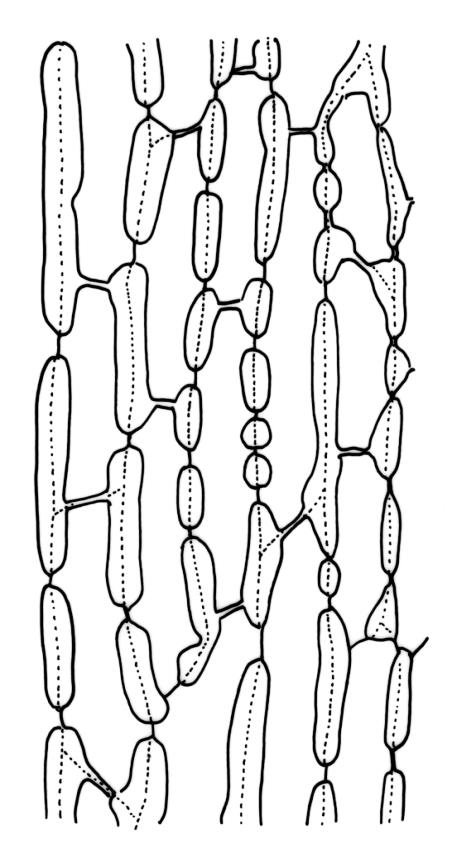
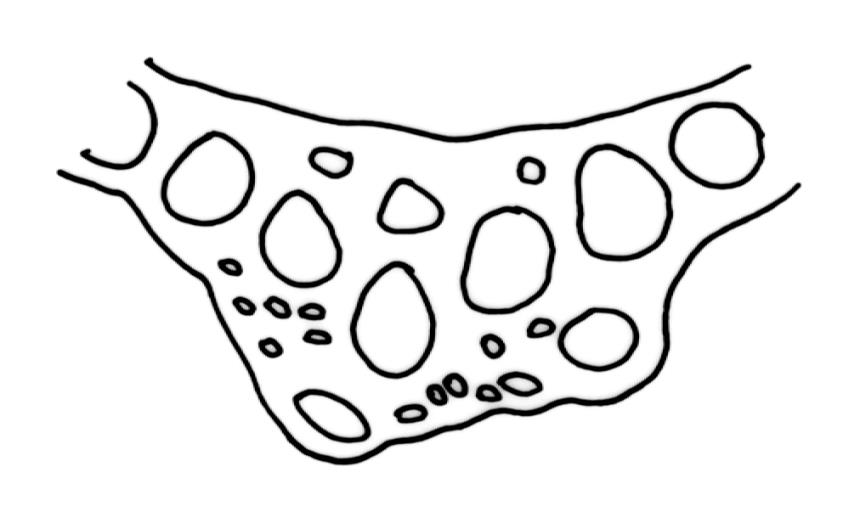
Plants yellow-green to golden when fresh, becoming yellow-brown when dry, forming dense cushions to at least 150 mm diam. Stems erect, aggregated and self-supporting, (30–)50–70(–90) mm, branched by forking, in cross-section with firm-walled cortical cells and a very weak central strand (in N.Z. material), beset with pale brown, smooth rhizoids. Leaves erect-appressed, both moist and dry, ovate-lanceolate and narrowly tapered to an acute or slightly rounded apex, unbordered, concave below, broadly tubulose above, entire or with a few weak teeth at extreme apex; plane or weakly inrolled at margins, (2.8–)3.5–5.8 × 1.1–1.3 mm (well-developed leaves under cover slip); mid laminal cells elongate, mostly rectangular-vermiculate, incrassate, strongly porose, mostly 48–78 × 9–12 µm, becoming more irregular near apex; alar cells partially or predominantly bistratose, forming a large orange-brown group, separated from the costa by a group of basal cells that are ± thin-walled, oblong-rectangular, non-porose, and mostly unpigmented. Costa 0.1–0.125 the width of the widest part of the leaf, percurrent, not toothed abaxially (in N.Z. material), at mid leaf with a central layer of (3–) 4–6 guide cells and two stereid bands (sometimes poorly developed, as in Plate 25, fig. D) and with a few large-lumened cells on the abaxial surface, in upper leaf cross-section rounded or weakly ridged abaxially.
Probably pseudautoicous. Sex organs and sporophytes not known from N.Z.
Sainsbury 1955, pl. 21, fig. 1 (as D. scoparium); Ireland 1982, pl. 89; Smith 2004, fig. 59, 4–6, 9; Hallingbäck et al. 2006, p. 343.
Confusion is most likely in N.Z. with entire-leaved forms of Dicranoloma robustum, but the presence of the unpigmented cells at the leaf base (absent in D. robustum) and the nature of the alar cells provide reliable distinctions. Such forms of D. robustum seem also to invariably have widely spreading leaves.
SI: Otago (St Bathans Range, Remarkable Range, Old Man Range, Old Woman Range, Rock & Pillar Range, Lammermoor Range, Garvie Range).
Apparently bipolar. Widespread in cool and oceanic regions of the northern hemisphere.
Forming dense cushions in cushion bogs, weak flushes, or boggy areas in tussock grasslands in subalpine and alpine areas of the Otago Ranges. Ranging in elevation from c. 980 m (Lammermoor Range) to at least 1700 m (Old Woman Range) elevation. Often associated with Oreobolus pectinatus, straight-leaved growth forms of Dicranoloma robustum, and Sphagnum cristatum. Polytrichum juniperinum, Straminergon stramineum, and Warnstorfia sarmentosa are less frequent associates.
Sainsbury (1955, p. 123) recorded D. scoparium from the Rock & Pillar Ra. (Otago L.D.), based on material collected by K.W. Allison; he reported this material as a "variety or form" of D. scoparium [D. scoparium var. orthophyllum Brid., Muscol. Recent. Suppl. 1: 173, 1806]. Although the name D. scoparium has been applied to Otago L.D. material since Sainsbury’s report (e.g., Fife 1995 and various workers in herb.), its application here is misleading. The Otago high-elevation material invariably has erect-appressed ovate-lanceolate leaves that are entire or nearly so, and with costae lacking both abaxial spines and ridges. Representative D. scoparium from the northern hemisphere has mostly secund and strongly serrate leaves with costae markedly ridged abaxially. On morphological grounds the Otago material is most similar to Dicranum leioneuron Kindb., a species with a Nova Scotian type. Smith (2004) considered it in Britain to be an occupant of "Sphagnum tussocks on raised and blanket bogs and in drier habitats under heather and in thin turf over boulders". Otago material compares very well morphologically to descriptions in Smith (2004) and in Ireland (1982), as well as to Newfoundland material. Our material occupies boggy sites not unlike those described for Europe.
The geographic isolation of the Otago populations of Dicranum could be used to argue that they represent an undescribed species. The recognition of a northern hemisphere species in N.Z. on morphological bases alone is certainly open to criticism, and molecular investigations could help to clarify the relationships of the Otago material. However, with present knowledge it is unjustified to describe the Otago material and it is deemed preferable to apply the Canadian name D. leioneuron to it, especially given the lack of representative (i.e. "typical") D. scoparium in N.Z.
Dicranum scoparium var. orthophyllum Brid. was recorded from mainland Australia (N.S.W.) by Streimann & Klazenga (2002), but no additional information about this record has been located. The south-east Asian D. psathyrum Klazenga [J. Hattori Bot. Lab. 87: 118, 1999] is a predominantly epiphytic species that, judging from Klazenga’s (1999) illustration, is certainly not conspecific with the Otago collections.
N.Z. material does not constantly produce flagelliferous branches, but these are present in some collections (e.g., J. Child 6032 from Old Man Range, CHR 429009). Flagelliferous branches are an inconstant feature of British populations of D. leioneuron (Smith 2004, p. 202). Neither sporophytes nor sex organs have been found in N.Z. collections and they are likewise unknown from Britain (Smith 2004). Hallingbäck et al. (2006, p. 343) report that dwarf males occur in Scandinavian material and that sporophytes are "relatively rare" in this species generally.
The bistratose nature of the alar cells in N.Z. material is often inconspicuous but, with care, can be observed under the compound microscope. A hand section of the alar region is readily accomplished and then the partially or predominantly bistratose nature of the alar group is obvious under either a stereoscope or compound microscope. The presence of a conspicuous group (not illustrated here) of non-pigmented, non-porose, and rather thin-walled cells between the alar group and the costa base is also a constant feature of N.Z. D. leioneuron.