Class
Family
Genus
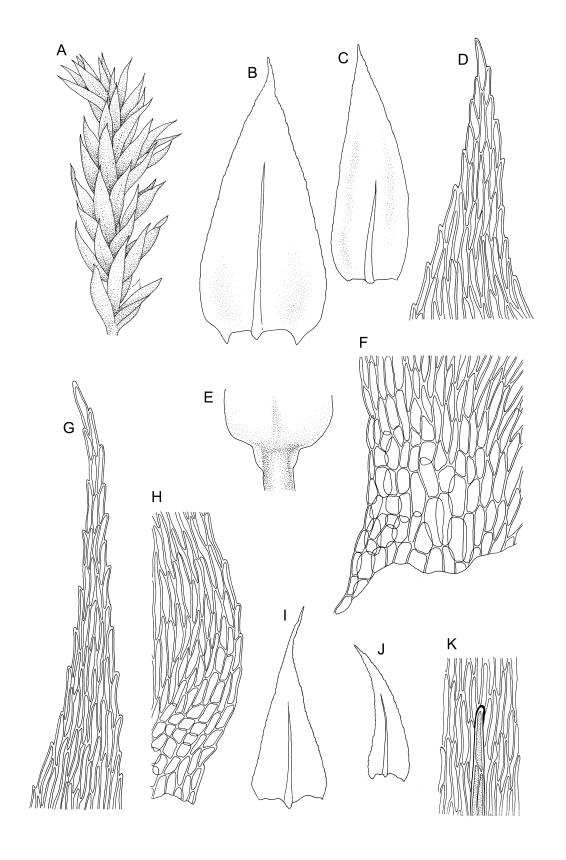
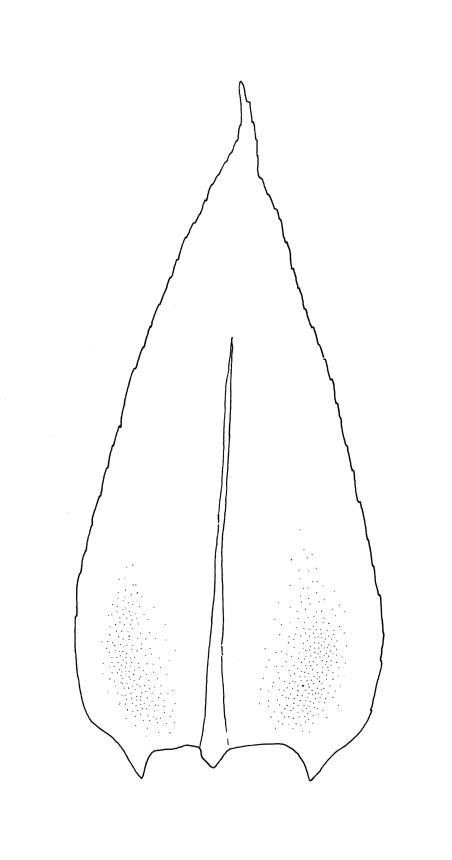
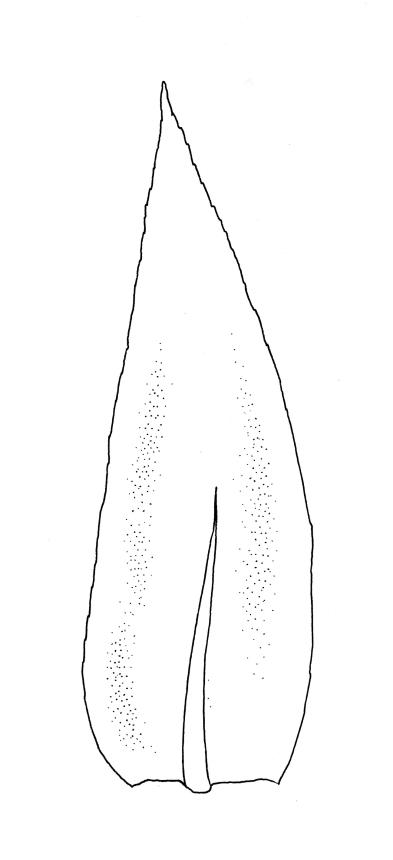
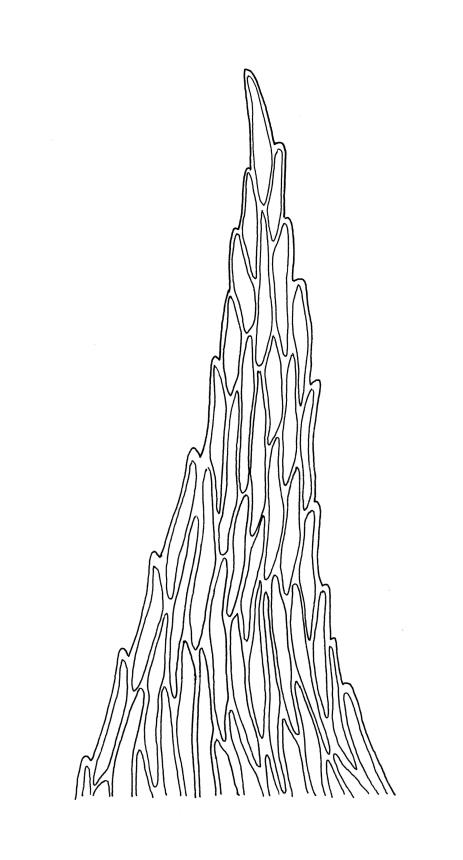
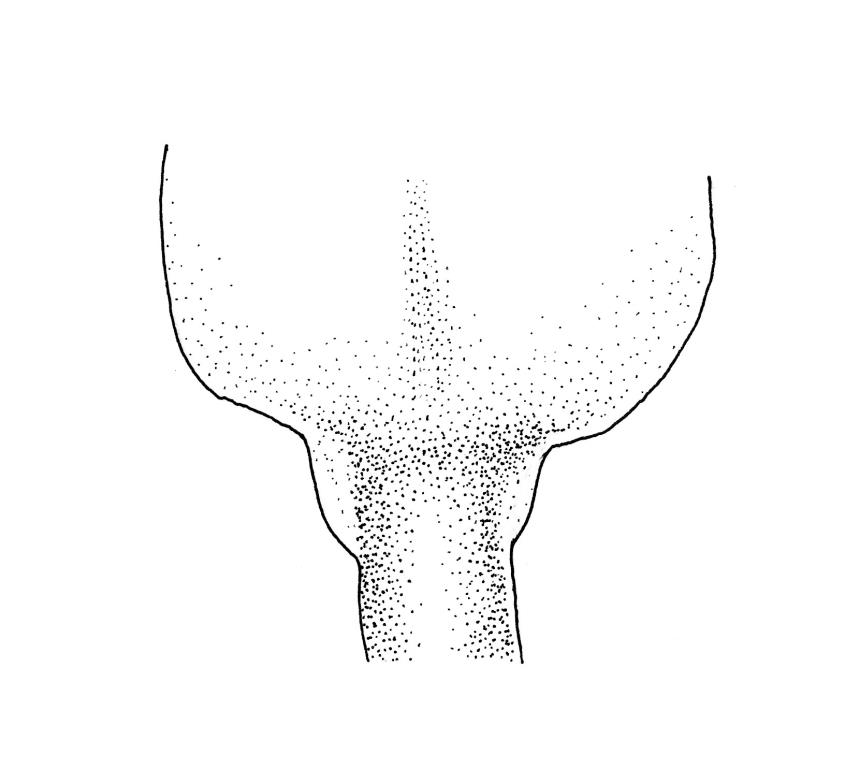
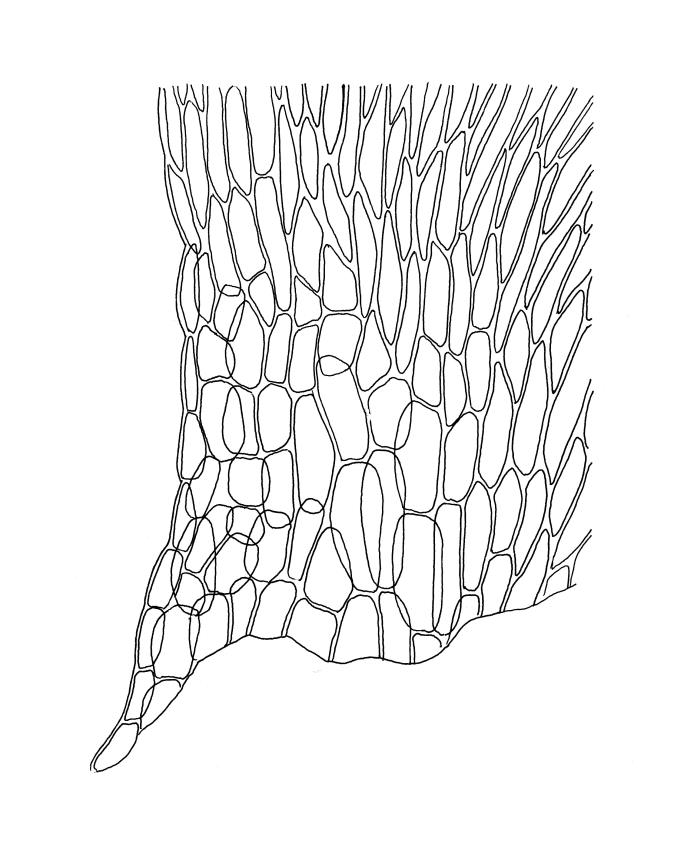
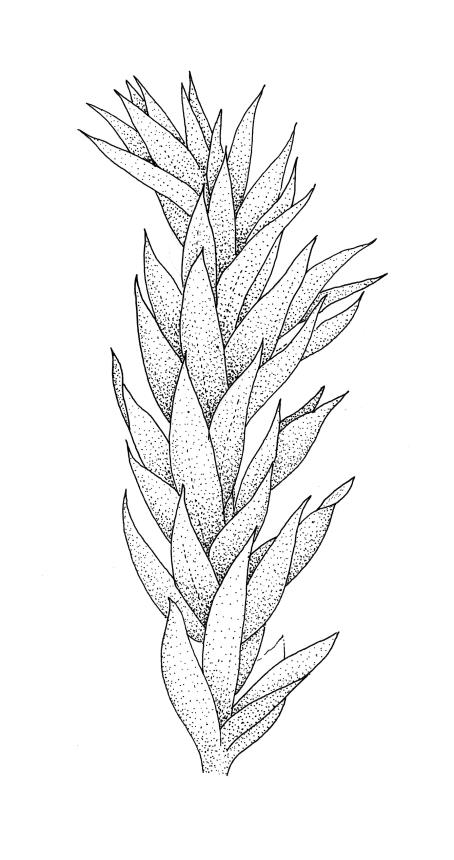
Plants medium-sized or robust, coarse, yellow-green or bright green, forming loose and untidy mats. Stems prostrate to ascendant and self-supporting, usually arching, irregularly but much branched, pale green to brown, to at least 80 mm long, in cross-section round or elliptic, with a distinct central strand and 3–4 layers of thick-walled cortical cells, with fascicles of pale brown, smooth rhizoids arising from abaxial side of leaf bases. Branches variable in length, commonly 5–20 mm long, with leaves slightly complanate, spreading. Stem leaves spreading, symmetric, not or only weakly plicate, scarcely altered when dry, ovate, acuminate, sometimes twisted at apices, plane, scarcely concave, decurrent, serrulate nearly to base, less often only above, with a large alar group, (2.0–)2.5–3 mm × c. 1 mm. Branch leaves smaller and narrower, 1.5–2.5 mm, slightly plicate when dry, with a smaller alar group, not or only weakly decurrent. Costa c. 30 µm wide (⅓ above insertion), extending c. ½–⅔ the length of the leaf, ± evenly tapered throughout, lacking a terminal spine. Upper laminal cells smooth, rather thin-walled, linear, c. 100–150 × 9 µm, basal cells shorter in 3–5 rows; alar cells (of stem leaves) in a large but ill-defined group that extends c. ⅔ from the margin to the costa and into the decurrency, inflated and rather thin-walled, oblong.
Autoicous or polygamous. Perichaetia scattered on stems, apparently enlarging after fertilisation; perichaetial leaves acuminate from an oblong base, ecostate, erect-spreading to squarrose. Perigonia ovoid, scattered on lower portion of stems, with ecostate, ovate, shortly acuminate, sometimes ± spreading bracts and filiform (7–9-celled) paraphyses. Setae (15–)20–30(–40) mm, scabrose throughout, weakly sinistrorse throughout, red-brown; capsules ± horizontal, asymmetric, curved, oblong-ovoid, c. 2.5–3.0 mm long (excluding operculum), dark yellow-brown; exothecial cells ± oblong, (24–)30–60(–75) µm, thick-walled. Annulus of 2–3 cell rows, falling with the operculum; operculum conic, acute and nearly black at apex. Exostome teeth lanceolate; endostome with well-developed nodose cilia in groups of 2–3. Calyptra c. 3 mm long. Spores 12–18 µm, finely papillose.
Scott & Stone 1976, pl. 83; Crum & Anderson 1981, fig. 516; Hedenäs 2002, fig. 5 c–d; Meagher & Fuhrer 2003, p. 52–53; Seppelt 2004, fig. 32.
Sterile B. rutabulum is sometimes confused with Rhynchostegium tenuifolium. Brachythecium rutabulum is a more robust species, with larger leaves (stem leaves c. 2.5–3.0 vs c. 1.6–2.0 long), differentiated alar cells, and decurrent stem leaves. Numerous sporophytic characters, including seta ornamentation and operculum form, also distinguish the two species.
Sterile material of Eurhynchium speciosum can also be confused here. Brachythecium rutabulum can be distinguished from E. speciosum by its more strongly dimorphic leaves, less strongly toothed margins, and the absence of an abaxial costal spicule.
Confusion can also occur with the more wiry, duller, and aquatic Platyhypnidium austrinum. Additional distinguishing features are discussed under that species. There appears to be little difficulty in distinguishing B. rutabulum from B. salebrosum in N.Z.
NI: N Auckland, including offshore islands (HC, LB, GB), S Auckland, Gisborne, Hawke’s Bay, Taranaki (near Ohura, Awakino Scenic Reserve), Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A; C; M.
More or less bipolar, but with a few scattered tropical localities. Tasmania*, mainland Australia*, North America*, Europe*, Asia. Reported by Crum & Anderson (1981) from Macaronesia, North Africa, Hawai‘i, Guatemala.
Weedy and occupying a wide range of substrates, including duff, humus, peat, sand, rocks (including limestone and concrete), and rotten logs in a wide range of vegetation types, including southern beech or mixed podocarp forest, exotic plantations, riparian Salix forest, scrub, and both native and modified grassland. Often in moist or even waterlogged situations such as seeps or stream/river margins, and shade tolerant. Only rarely epiphytic (and then probably subject to flooding). On the North I. occurring from near sea level to at least 740 m (Erua, Wellington L.D.), and on the South I. occurring from near sea level (several localities) to at least 1675 m (Pisa Range, Otago L.D.) elevation.
Brachythecium rutabulum is both weedy and highly variable, ranging from compact, stunted forms with stems less than 20 mm, to robust, openly branched forms with stems in excess of 80 mm. The latter are more representative. In compact forms, which are usually associated with drier situations, the stem leaves can be quite crowded and their decurrencies less conspicuous than normal. Microscopic features, especially the enlarged and oblong alar cells, however, are diagnostic even in these plants. The distinctly dimorphic, non-plicate leaves, decurrent stem leaves, and well-differentiated alar cells usually create a gestalt that permits recognition of this weedy species despite its variability.
Smith (2004) described this species in Britain as “a fast-growing species, especially of damp woodland in base-rich situations” and considered it to be “rare or absent on acidic nutrient-poor substrates.” Both he and Crum & Anderson (1981) reported a wide range of haploid chromosome numbers from different parts of its wide range.
Brachythecium rutabulum often grows at stream margins and sometimes occurs as an emergent in small, swift streams. Such material invites comparison, if only by virtue of its habitat, to B. rivulare, a northern hemisphere species that has been recorded from both Tasmania and mainland Australia (Hedenäs 2002). No N.Z. material is accepted as B. rivulare. The aquatic forms noted above have stem and branch leaves serrulate ± to their base, a tendency for leaf apices to become twisted when dry, and generally less strongly differentiated alar cells than those occurring in B. rivulare.
Dixon (1929, p. 322) recorded what he termed “B. rutabulum var. robustum Bryol. Eur.” from the Mt Cook district, citing a collection by J. Murray. Dixon’s record presumably formed the basis of the N.Z. citation for this taxon in Wijk et al. (1959). The Murray specimen has not been examined. Dixon’s differentia for this alleged variety (quoted by Sainsbury 1955) do not agree with those given by Limpricht (1895–1903, p. 109). Many infra-specific taxa recognised by earlier workers within B. rutabulum, including this one, no longer enjoy wide acceptance (e.g., Corley et al. 1981; Smith 2004) in European literature. For these reasons, plus the variability of B. rutabulum in N.Z., the so-called var. robustum is not discussed further.