Class
Family
Genus
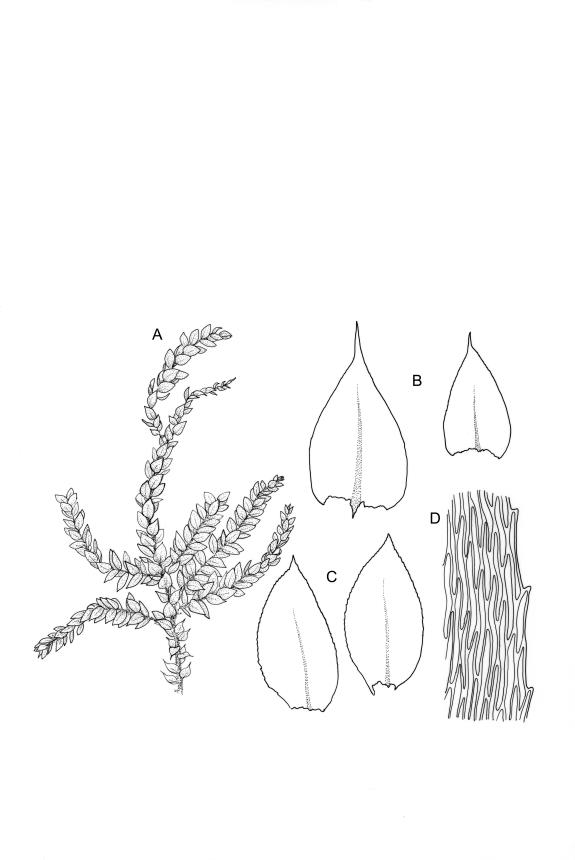
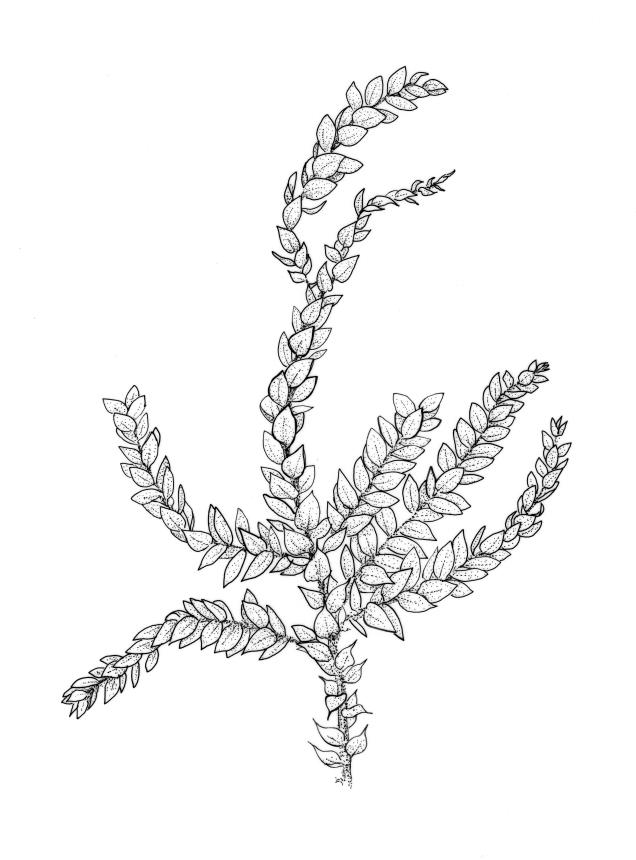
Plants medium-sized, yellow- or dirty-green, lustrous, forming rather rough and irregular wefts. Stems creeping, irregularly to subpinnately branched, short or elongate (to >100 mm long); branches ascendant, simple or sparsely branched, mostly 6–12 mm. Stem leaves rather distant, spreading, broadly ovate, abruptly tapered to a short and sometimes twisted acumen, sharply serrulate throughout, weakly decurrent, scarcely concave, not plicate, 1.5–1.8 × 0.65–1.0 mm; costa tapered, extending c. ¾ the leaf length, lacking a terminal spine; mid laminal cells linear, firm-walled, weakly prorate at upper end, mostly 120–165 × c. 6 µm, becoming shorter and broader at insertion; cells of the acumen not differentiated; alar cells scarcely differentiated, forming weak decurrencies. Branch leaves differentiated, complanate, broadly ovate to nearly elliptic, acute or broadly acuminate, weakly asymmetric at base, c. 1.4 × 0.6 mm; costa extending c. ¾ to ⅞ the leaf length, nearly uniform in width throughout, with a conspicuous terminal abaxial spine.
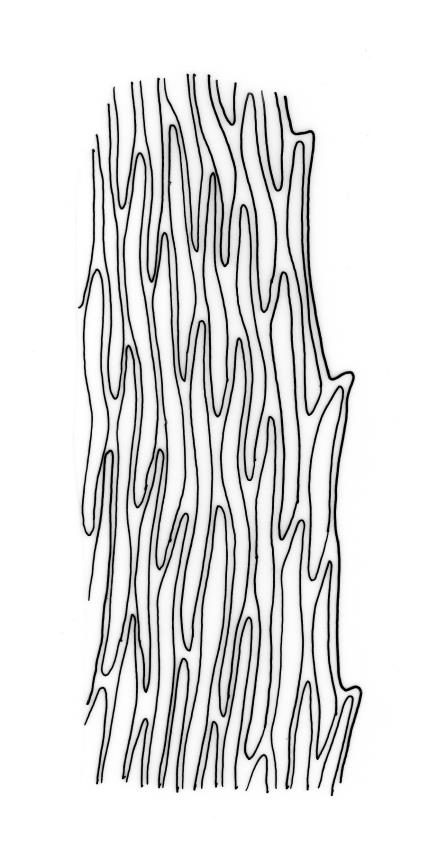
Autoicous or synoicous. Perichaetia scattered on stems, with leaves squarrose from a sheathing base, ecostate. Perigonia, scattered if present, with wide-spreading bracts and filiform paraphyses. Setae slender and elongate, 15–22 mm, red-brown, finely papillose throughout (usually not visible under hand-lens); capsules oblong-cylindric, narrowed below mouth when dry, ± horizontal, c. 2.0 mm; exothecial cells mostly rounded-oblong, thick-walled, very weakly thickened in corners; stomata few at capsule base; annulus and operculum not seen. Exostome teeth as for genus, c. 500 µm; endostome as for genus, cilia paired. Spores 18–21 µm, smooth.
Smith 2004, fig. 287, 1–4 (as Oxyrrhynchium speciosum).
K; NI: N Auckland, S Auckland. SI: Westland? (see notes below)
Probably adventive. Largely confined to Europe but reported from Iran, Saudi Arabia, China and Macaronesia by Smith (2004). Jessica Beever (pers. comm., 3 May 2019) considers this species to be “quite common” in N Auckland L.D. The tentative Westland record is from the “Lake Brunner shore” and collected by M.F. Sinclair in 1970. The small sterile fragment I have seen of this collection appears to be correctly determined and bears a note by K.W. Allison that he had seen one sporophyte with a “sparsely and lowly papillose” seta. The occurrence of this species on the South I. requires confirmation.
Occurring on Kermadec Islands from sea level to c. 150 m; on the North I. from near sea level to 450 m (Whirinaki Forest, S Auckland L.D.). On Raoul I. (Kermadecs) this species is often collected in a variety of mostly modified vegetation types, including underneath Mysore thorn (Caesalpinia decapetala); it is often associated with Racopilum cuspidigerum var. convolutaceum. The majority of the North I. collections are from either damp hollows or damp sites adjacent to streams and probably subject to periodic flooding. There is little constancy in associated vegetation types; they include native forest (with kahikatea, taraire, etc.), mixed bracken and Phormium, and adventive grasses. The indicated substrates were soil and rotten logs.
Beever et al. (1996) discussed the typification of the synonymous Hypnum trachypelma Müll.Hal. Material reported by Sykes (1977) from the Kermadecs as Rhynchostegiella vitiensis Dixon [Proc. Linn. Soc. New South Wales 55: 300, 1930] was tentatively referred to E. speciosum by Beever et al. (1996). There are three sterile specimens in herb. J.E. Beever from N Auckland L.D. that have branch leaves more lanceolate and plicate when dry than the other N.Z. material.
The relationship between the present species and the widely distributed northern hemisphere E. hians (Hedw.) Sande Lac. [Ann. Mus. Bot. Lugduno-Batavi 2: 299, 1866] is unclear and beyond the scope of this eFlora treatment. All fruiting N.Z. material is either synoicous or autoicous (or polyoicous?) and thus conforms with the sexuality of E. speciosum given in European floras (cf. Limpricht 1895–1903; Smith 2004). There are, however, several sterile N.Z. collections that are probably dioicous (which is the sexual state of E. hians in central Europe, fide Limpricht 1895–1903) and eastern North America (fide Crum & Anderson 1981). I can find no other morphological differences between the limited North American material of E. hians studied (including S. Rapp 61, CHR 649698) and confirmed NZ material of E. speciosum.
No comparison has been made of E. speciosum to E. laevisetum Geh. of eastern Australia. Apart from alleged differences in sexuality, the dioicous E. laevisetum is distinguished from the present species by Hedenäs (2002) by a series of overlapping quantitative characters that are not fully convincing.