Class
Family
Genus
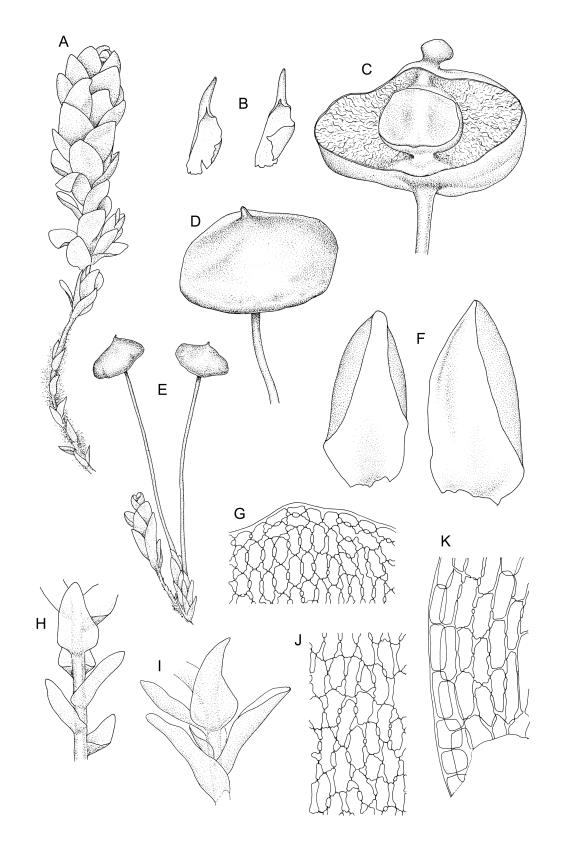
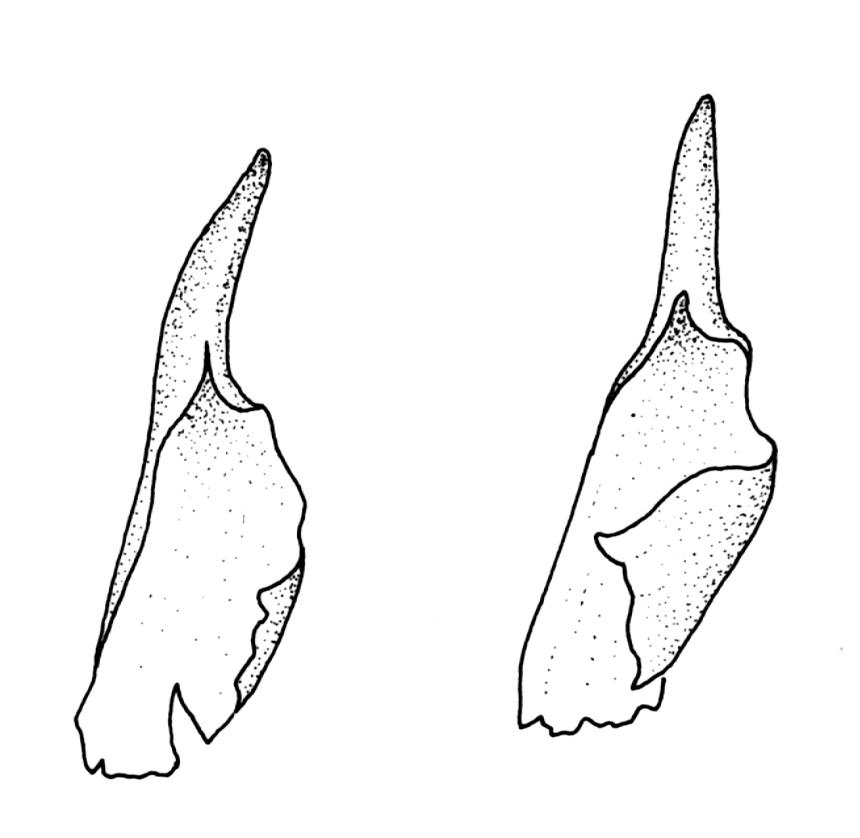
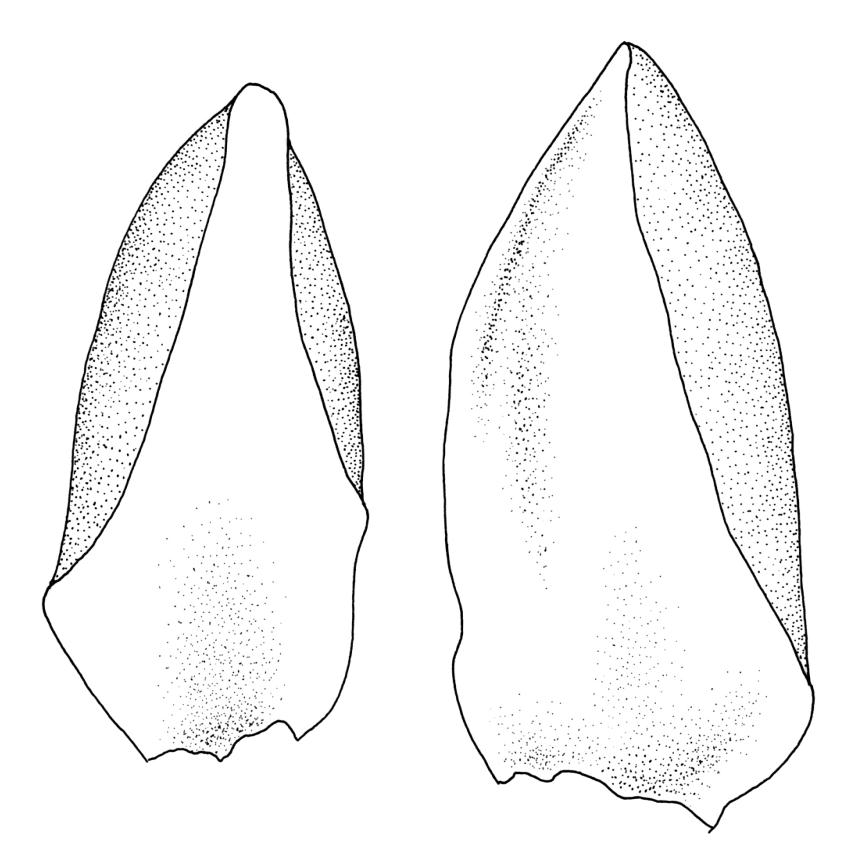
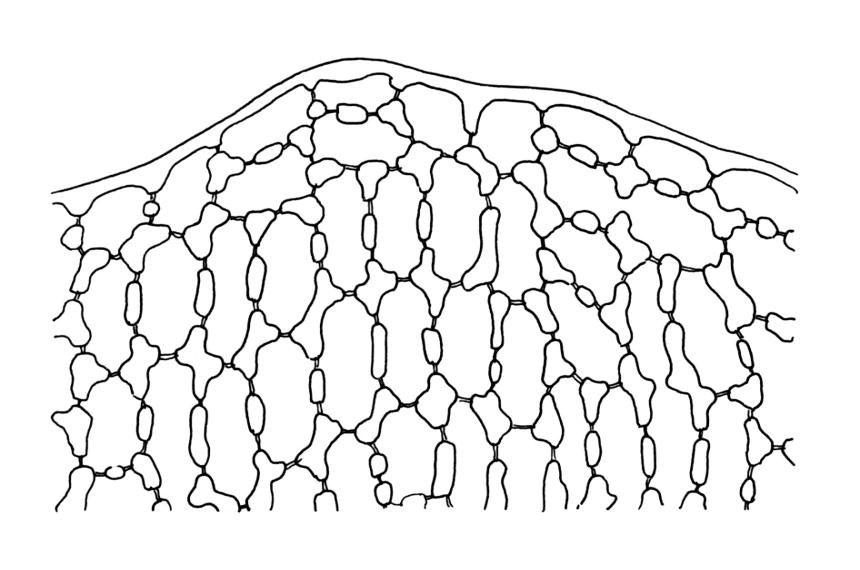
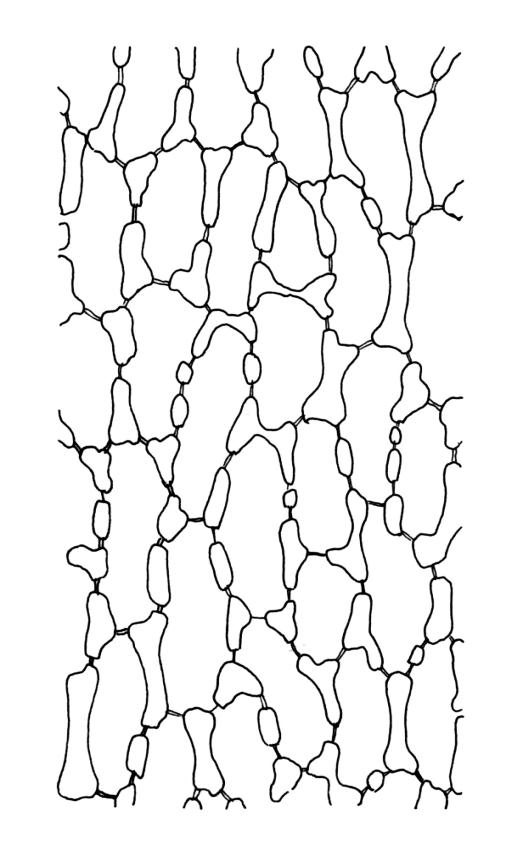
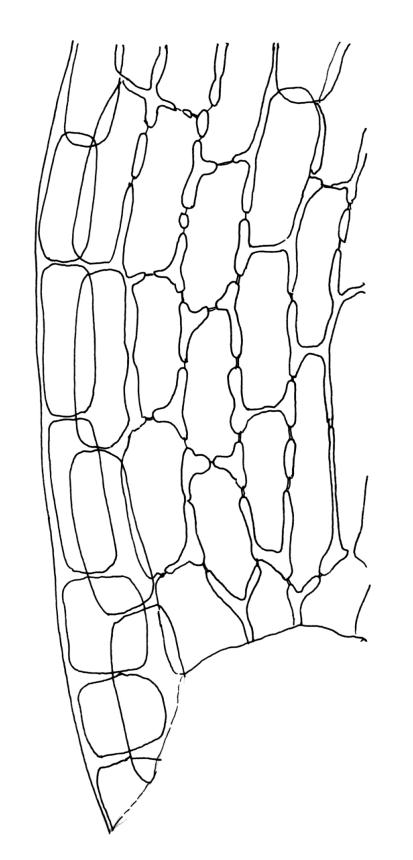
Plants strongly julaceous, with dark brown, creeping, and subterranean primary stems. Secondary stems to c. 40(–60) mm in sterile material, usually <20 mm when fertile, variably sheathed by a weft of pale, smooth rhizoids (originating from outer cortical cells) with oblique end walls. Leaves broadly elliptic to nearly cochleariform, rounded and non-reflexed at apices, mostly 2.0–3.2(–3.5) × 1.4–1.8(–2.0) mm, strongly concave, mostly loosely imbricate both moist and dry, sometimes erect-spreading in robust sterile plants, often funiculate in sinistrorse ranks on sterile shoots, narrowly and obscurely decurrent; margins erect throughout or weakly and narrowly reflexed near base; mid laminal cells mostly oblong but somewhat variable in shape, very thick-walled and highly porose, strongly opaque, mostly c. 45–72 × 15–18 µm, becoming shorter and broadly rhombic to subquadrate near leaf apex; basal cells long-rectangular to long-hexagonal, to c. 150 µm, thick-walled and porose; marginal cells regularly oblong in a few rows; alar cells shorter and wider than adjacent laminal cells, sometimes weakly auriculate; cells of decurrencies ± quadrate, usually not stripping off with leaves.
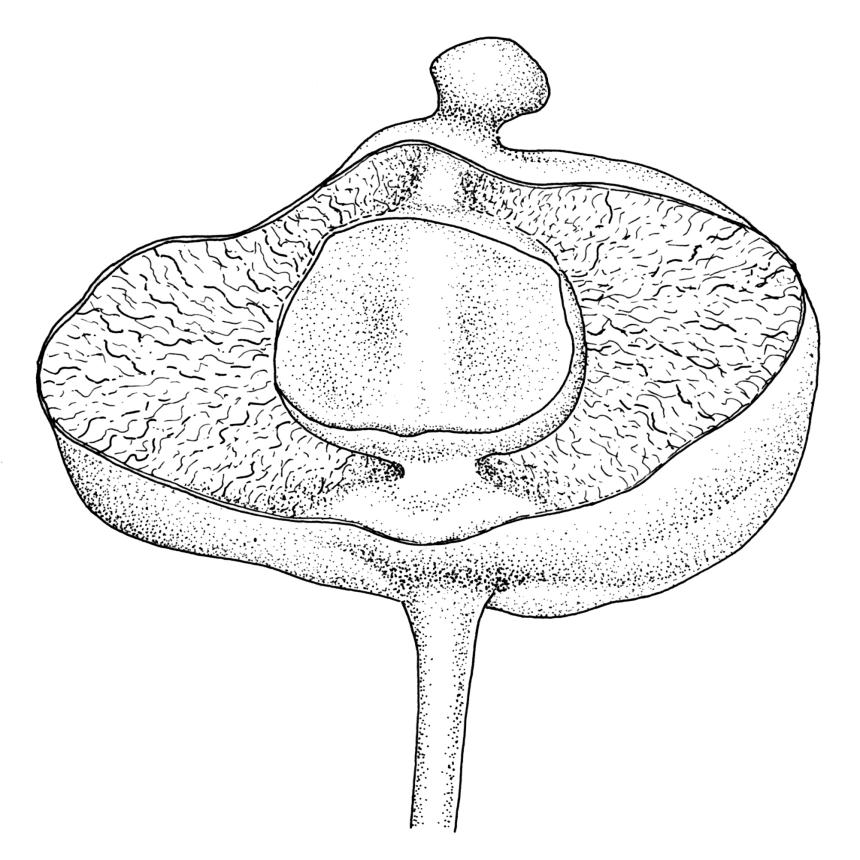
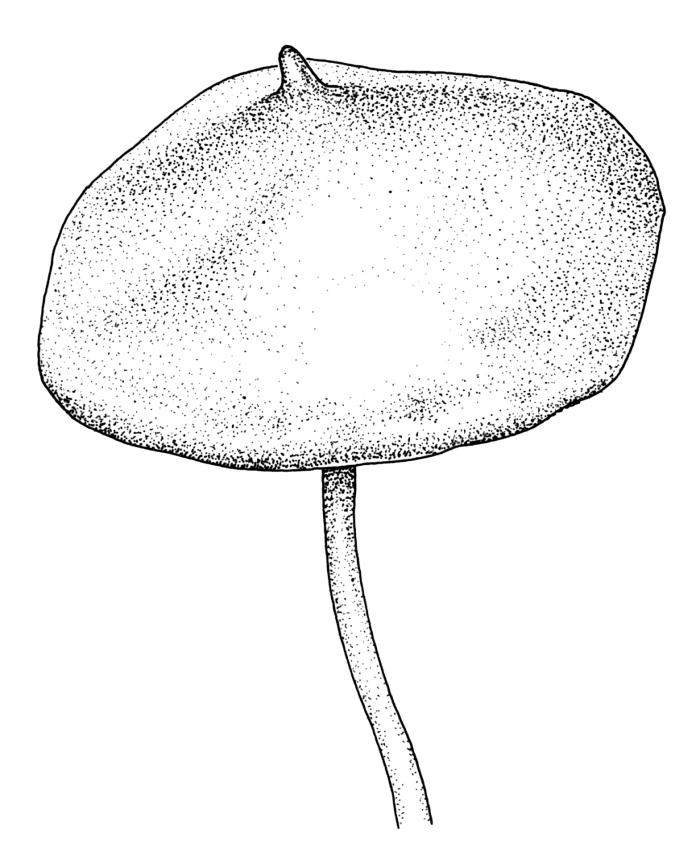
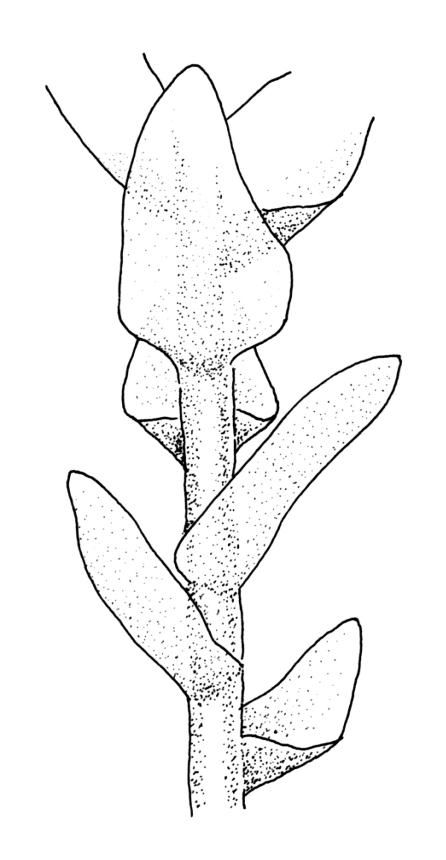
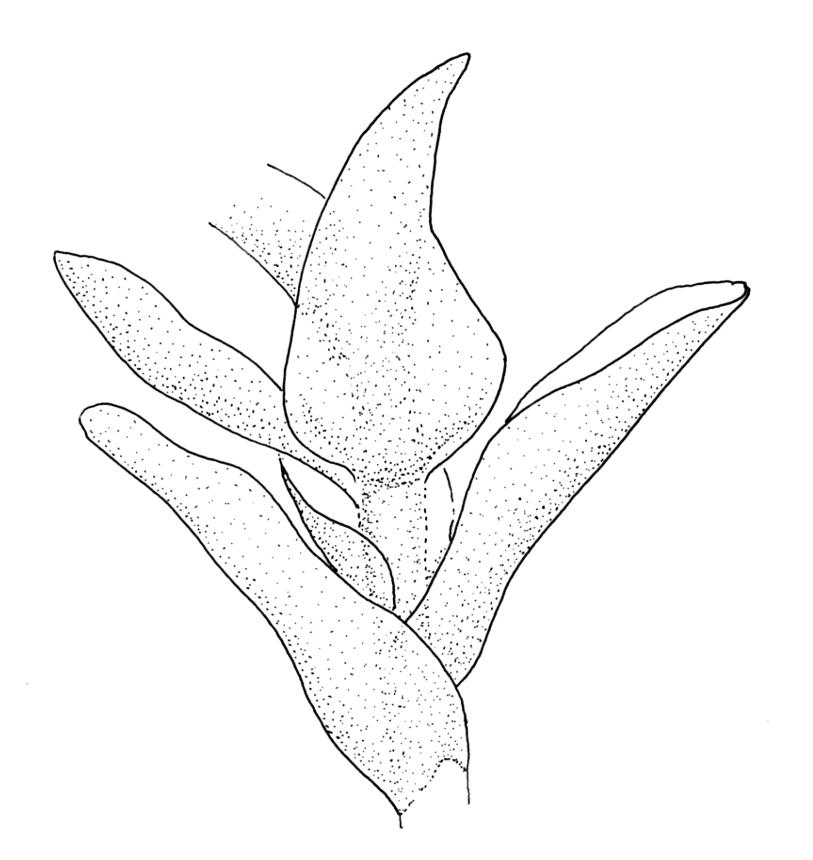
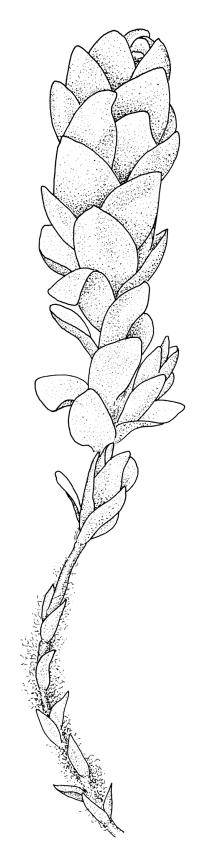
Perichaetia before fertilisation c. 1.5–2.0 mm long, with leaves widely spreading and surrounding several archegonia and filiform paraphyses, following fertilisation expanding to c. 7 mm long, with leaves broadly oblong-elliptic, obtuse, to 4.5 mm, not or very weakly reflexed at apex, sheathing the lowermost 2–3 mm of seta. Perigonia sessile, scattered and lateral on secondary stems, gemmiform, with very broadly ovate, obtuse, and highly concave bracts c. 0.8 mm long enclosing the numerous antheridia and no paraphyses. Setae 10–35 mm, pale yellow; capsules globose when immature, becoming discoid at maturity (3.5–)6–12 mm diameter × c. 3 mm, bluntly rostrate (rostrum 0.6–0.9 mm, rarely swollen apically) at apex, bright red-orange when fresh, fading to pale or dirty yellow with age; spore sac as per genus. Calyptra as per genus. Spores oval, ± smooth, 33–45 µm.
Malcolm & Malcolm 2003, p. 52 (as P. grandiglobum var. decurrens); Fife & Dalton 2005, fig. 2, B–E; Malcolm & Malcolm 2006, p. 31, 83, 84, 202, 237, 246.
SI: Nelson, Canterbury (Bealey Glacier Track), Westland, Southland (Lake Hankinson, Percy Saddle). The bulk of the known localities are in Nelson and Westland L.D. Nearly all collections with sporophytes are from a restricted area encompassing the general vicinity of Westport and Greymouth, the Stockton and Denniston plateaux, and the Paparoa Range. A paucity of collections from south of the latitude of Ōkārito (c. 43°15' S) may be a collection artefact. Material of uncertain identity from St is discussed below.
Endemic.
At pool margins in cushion bogs, pākihi (a nutrient-depleted swamp with highly podsolised hard-pan soils), and on sandstone pavements (as on Denniston Plateau, Nelson L.D.); also in wet sites in subalpine scrub and red tussock grassland including associated flushes. Occasionally submerged. Frequently associated mosses include Breutelia elongata, Campylopus introflexus, Dicranoloma robustum, Ptychomnion aciculare, Rhacocarpus purpurascens, Sphagnum cristatum and S. falcatulum, while associated hepatics include Anastrophyllum schismoides, Isotachis spp., Kurzia helophila, Riccardia australis, Temnoma pulchellum, and Trichocolea mollissima. Pākihi sites where it occurs are usually characterised by scattered small trees of manuka (Leptospermum scoparium). P. ovalifolium frequently (as on Denniston Plateau) grows in close proximity to the restiad Empodisma minus, the sedge Schoenus pauciflorus, the ferns Gleichenia spp., and sundews (especially Drosera spathulata). Other often associated vascular plants include Donatia novae-zelandiae, Euphrasia disperma, Schizaea australis, and Utricularia monanthos. Ranging from c. 50 (near Westport) to 1430 m (Cobb Valley, both in Nelson L.D.) and known only from west of, or very close to, the main divide on the South I.
Many collections from the Denniston Plateau-Stockton Plateau-Westport vicinity in Nelson L.D. are fertile, whereas capsules from outside this general vicinity are less common. Capsules mature in spring to late summer. The globose immature capsule becomes strongly flattened, discoid, and bright red-orange to scarlet with maturity (see Fife & Dalton 2005, fig. 2). The spore sac in a mature capsule of 6 mm diameter is c. 2.2 mm diameter (in M.J.A. Simpson 8561, CHR 351331); the spore sac is attached to the exothecial wall by anastomosing cellular filaments. Plants with mature capsules are completely unmistakeable and, when fruiting, this is arguably the most striking moss in the N.Z. flora. The strongly julaceous shoots and ± cochleariform, ecostate leaves and very thick and strongly porose laminal cell walls are also highly characteristic.
The means by which the spores are released from the globose capsules are unknown, although in mature material the exothecial wall sometimes disintegrates, possibly following mechanical disruption, and peels away from the still intact spore sac. Developing sporophytes probably require more than a single growing season to fully mature. In the wet sites where Pleurophascum ovalifolium characteristically occurs, the large air spaces within the capsule might facilitate dispersal by water. No internal spore germination has been observed.
Repeated efforts to find perigonia in fruiting specimens have failed. The pale, nearly white, filaments that often sheath the secondary stems arise from outer cortical cells of the stem and have oblique end walls; they are interpreted here as rhizoids rather than fungal hyphae.
Sainsbury (1952) originally described this taxon as a variety of the Tasmanian P. grandiglobum but subsequently (1955, p. 141) suggested that it "may well prove to be entitled to specific rank". In P. grandiglobum, the upper leaves of fertile plants have reflexed or narrowly recurved upper margins and strongly reflexed apices. Capsules in P. grandiglobum are consistently globose and dull brown-orange or grey-brown at maturity and have an apical rostrum that is shorter (0.2–0.35 mm) than that of P. ovalifolium. Other features distinguishing between the two species are detailed by Fife & Dalton (2005).
Two sterile fragments of Pleurophascum have been seen from Stewart I. (D. Glenny 9330b from Magog, Fraser Peaks area, CHR 571408 and C.D. Meurk & H.D. Wilson s.n., 5 Mar. 1980, from near South Cape, CHR 574228). In both collections leaves with weakly mucronate leaf apices and slightly recurved upper margins occur, raising the possibility that Stewart I. populations might differ from South I. populations. The collection of well-developed material from Stewart I. is required to clarify the status of these populations.