Class
Family
Genus
Orthotrichum alpestre sensu Sainsbury 1955
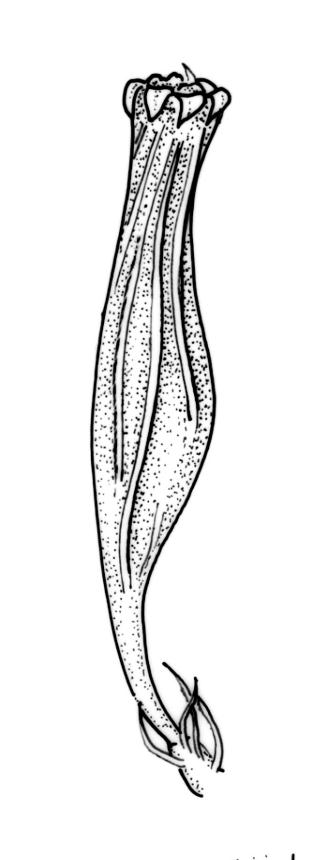
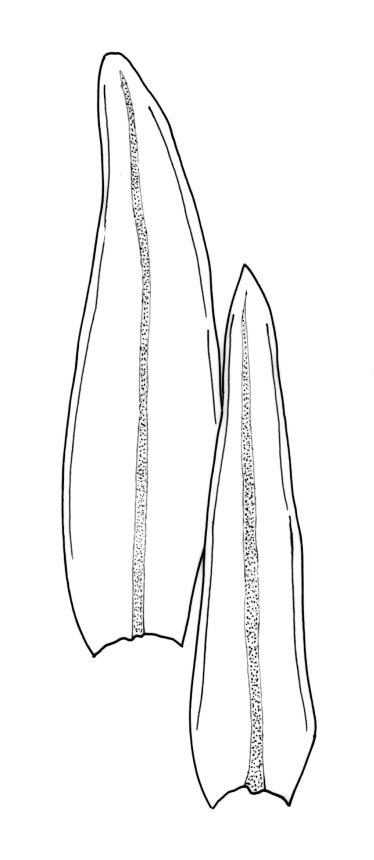
Plants forming loose or dense tufts, yellow-green to olive-green above, light brown to almost black below. Stems branched by innovation, 3.5–12 mm, with rhizoids only near the base. Leaves erect and slightly flexuose when dry, erect-spreading when moist, ovate- to oblong-lanceolate, acute, rounded-obtuse, or acuminate at apex, not plicate, finely and irregularly toothed or entire at apex, recurved from near base to near apex, (1.5–)1.8–3.5(–4.1) × 0.3–0.8(–1.2) mm; mid and upper laminal cells thick-walled, ± isodiametric, often ± oblate near margins, 6.5–15(–21) × 6.5–15(–20) µm, with 2–4 low, inconspicuous, and mostly unbranched papillae; basal laminal cells rectangular, 15–90 × 9.5–25 µm, usually thin-walled and lacking pores, but sometimes with thicker walls and porose, c. 6 rows at margins short rectangular and with thickened transverse walls. Costa as per genus.
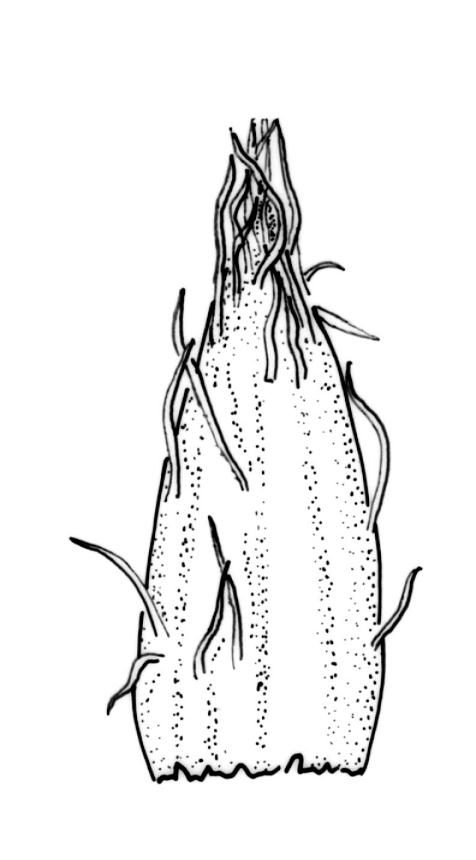
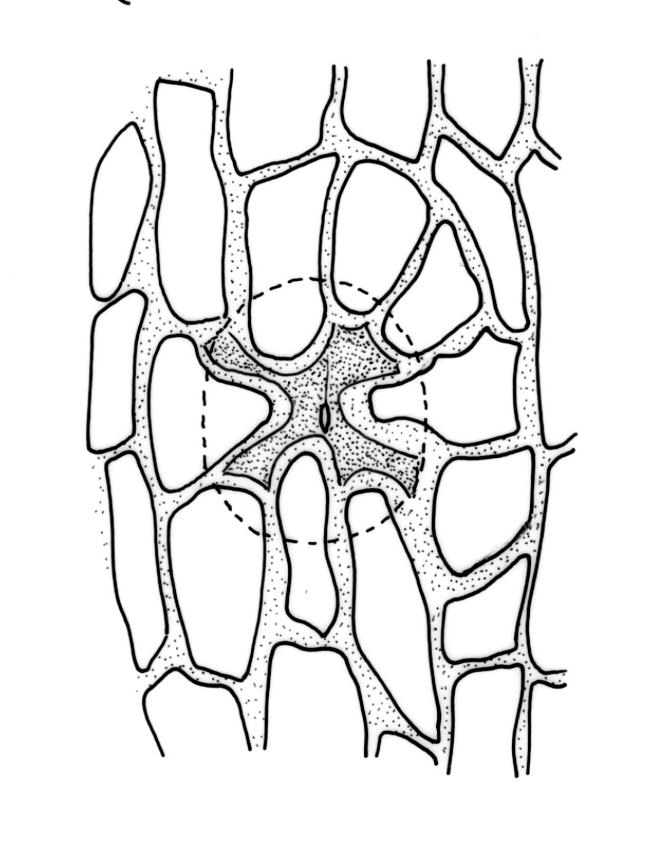
Gonioautoicous. Perichaetial leaves not differentiated. Perigonia gemmiform, below the perichaetia. Setae c. 2 mm; capsules immersed, emergent, or weakly exserted, narrowly cylindric, with 8 deep furrows nearly the full capsule length and constricted below the mouth when dry, less conspicuously furrowed when moist, 1 per perichaetium; exothecial cells differentiated into 8 yellow bands, each c. 4 cells wide, striate on surface; cells within bands rectangular with thick longitudinal and thin transverse walls; cells between bands rounded-elongate; stomata deeply immersed, restricted to mid and lower urn, the surrounding ("subsidiary") cells strongly protruding, but not completely covering the guard cells; operculum as per genus. Peristome double; exostome teeth 8, yellow, recurved when dry, finely and evenly papillose, with the papillae sometimes confluent in lines near the apex; preperistome absent; endostome segments 8, narrow and uniseriate (except near the base where sometimes 2 cells wide), yellow or hyaline, granulated or smooth near the base or smooth throughout. Calyptra campanulate-mitrate, yellow to ± brown, mostly red-brown at apex, somewhat plicate and split near the base, with many or few, yellow or hyaline, smooth hairs. Spores globose, isosporous and 1-celled, 16–21 µm diam., papillose, yellow-brown.
Lewinsky 1984, figs 32–33 (as O. longithecum); Lewinsky 1985, figs 35–47.
Orthotrichum assimile is recognised partly by its emergent and strongly 8-furrowed capsules with recurved peristome teeth. The leaves are erect and appressed when dry, in sharp contrast to the contorted leaves of O. calvum, the other predominantly epiphytic species with immersed stomata. The nature of the endostome and the degree of hairiness of the calyptra is somewhat variable. I have not seen gemmae ("fusiform or cylindric, 5–7 cells long"), which Lewinsky described as sometimes present on leaves.
Confusion (in sterile or poor material) can occur between O. assimile and O. cyathiforme; material of O. assimile has more acute leaf apices than does O. cyathiforme, but this difference alone is not reliable. In my opinion, observation of either stomata (deeply immersed in O. assimile vs superficial in O. cyathiforme) or the nature of the calyptrae (hairy in O. assimile vs naked in O. cyathiforme) is required for the confident differentiation of these two superficially similar species.
NI: N Auckland (near Waimate North), S Auckland (near Murupara), Hawke’s Bay (Otūpae, Sentry Box Scenic Reserve), Wellington (near Taihape, Masterton); SI: Marlborough, Canterbury, Otago, Southland; St.
Austral. Australia*. Reported from numerous localities in Chile and Argentina by Lewinsky (1985). Lewinsky (1984) characterised the N.Z. distribution of this species as "local in the central part of the North Island, common in the eastern and southern part of the South Island" and provided a detailed distribution map.
Primarily epiphytic, but rarely on non-calcareous rock. The majority of collections come from Salix and Populus bark. Less commonly occurring on the bark of a wide range of indigenous woody plants as well as adventives including Fraxinus, Sambucus, Ulmus, and "old fruit trees" (see Lewinsky 1984, tab. 1). Occurring from near sea level to 800 m.
According to Lewinsky (1984), Dixon (1926) overlooked the immersed (cryptopore) nature of the stomata when he erroneously placed O. longithecum R.Br.bis in the synonymy of O. hortense Bosw. (which has superficial stomata). Many workers after Dixon, including Sainsbury (1955), have treated the N.Z. and Australian material of O. assimile as conspecific with the northern hemisphere O. alpestre, but Lewinsky (1984, p. 440) detailed several morphological and ecological features that distinguish O. assimile from that widespread northern hemisphere species. She also noted a relationship between O. assimile (cited as O. longithecum) and the northern hemisphere O. pumilum Sw. In her publication dealing with South American representatives, Lewinsky (1985, p. 75) placed the N.Z. O. longithecum in synonymy with the Chilean and Argentinian O. assimile.