Class
Family
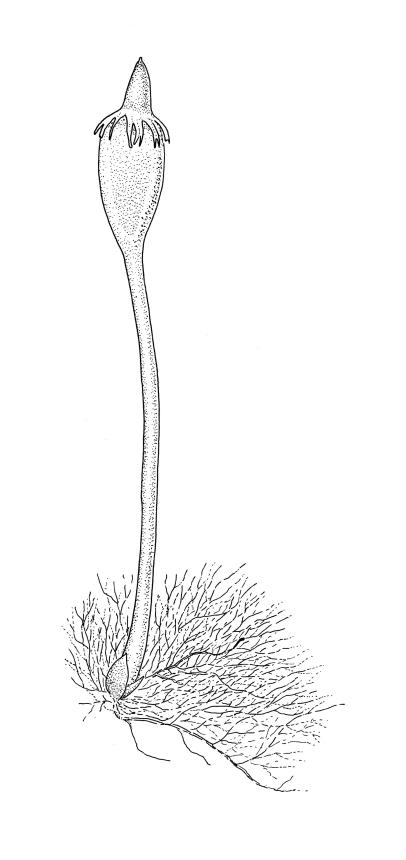
Plants minute, yellow- or red-brown, forming epiphytic mats on twigs or leaves. Protonema persistent, forming dense wefts; primary protonema with oblique walls, with or without fan-shaped holdfasts; secondary protonema erect, branching dichotomously, with transverse walls, with or without elongate terminal assimilative filaments. Shoots reduced to minute ♂ and ♀ buds arising from the primary protonema. Vegetative leaves absent.
Dioicous or autoicous (with ♂ and ♀ buds arising from same primary protonema). Perigonia and perichaetia reduced to minute, globose or ellipsoid buds arising from the primary protonema. Setae straight or nearly so, smooth; capsules small, erect or nearly so, campanulate or oblong-turbinate, with a distinct sterile neck; exothecial cells with or without thickened transverse walls, collenchymatous (in N.Z. species) or not; stomata present or absent, cryptopore or phaneropore; annulus poorly differentiated; operculum rostrate from a conic base. Peristome double; exostome teeth 16, linear-lanceolate, yellow-brown, outer surface transversely striate, inner surface with numerous low trabeculae; endostome with a basal membrane; endostome segments present or lacking; cilia lacking. Calyptra mitrate, smooth above, finely laciniate at base. Spores oblong or fusiform, unicellular or multicellular with transverse septa, green, smooth.
A genus of two species, one in Malesia and tropical south-east Asia and the other in Australasia. It is renowned for its highly simplified gametophytes, which are associated with morphologically complex sporophytes, and for its remarkably small size. The genus has attracted a high degree of morphological interest, particularly by German morphologists of the late 19th and early 20th centuries (Goebel 1892; Fleischer 1899; 1908; 1929; Renner 1935), since its type species was collected in Java in the 1890s.
A second wave of interest in the genus was provoked by K.W. Allison’s discovery in 1928 of the plant that was eventually named as E. trentepohlioides on the volcanic plateau near Ātiamuri, South Auckland L.D. (Dixon 1928; Renner 1935; Sainsbury 1951).
The generic placement of the N.Z. species treated here as an Ephemeropsis is problematic; this species has been plausibly placed in a segregate genus Archephemeropsis by Renner (1935), who argued (p. 88) that the superficial similarity of the two taxa is a result of convergent reduction in a hookeriaceous ("Hookeriaceen") group. Goffinet et al. (2009) recognised only the genus Ephemeropsis, which they place in the Daltoniaceae. The appropriate generic treatment cannot be resolved using morphological characters alone and needs to be considered in the context of the Daltoniaceae as a whole, probably using morphological and molecular or other non-traditional methods. For now it seems preferable to follow the generic assignment used by Sainsbury (1955) and hence most familiar to Australasian workers. The following morphological observations highlight the differences between the Australasian E. trentepohlioides and the Malesian E. tjibodensis.
The sporophytes of the Australasian species are more common than, and differ by many features from, those of the Malesian E. tjibodensis. The capsules of E. trentepohlioides, while longer (0.5–0.7 mm) than those of the Malesian species (c. 0.3–0.5 mm), have peristomes and stomata that are more difficult to observe microscopically. Exothecial cells in E. trentepohlioides have thin walls with collenchymatous thickenings while in E. tjibodensis exothecial cells have thickened and curved transverse walls and lack thickened corners. The peristome teeth of E. trentepohlioides are c. 150 μm long, not furrowed, strongly reflexed when dry, and have a median zigzag line extending either c. ⅓ the tooth length or absent altogether. By contrast the teeth of E. tjibodensis are c. 210 µm long, erect when dry, furrowed for c. ⅔ of their length and have a median zig-zag line extending nearly to the apex. In transverse section a peristome tooth of E. trentepohlioides has the inner layer thicker than the outer layer (c. 18 µm thick vs. c. 3–4 µm thick), in contrast to that of E. tjibodensis where the teeth have inner and outer layers of approximately equal thickness. In E. trentepohlioides, the inner layer in surface view is roughly equal in width to the outer layer at mid tooth, while in E. tjibodensis the inner layer is considerably narrower than the outer layer at mid tooth. In E. trentepohlioides, the endostomal basal membrane is highly reduced or absent with nil segments, while in E. tjibodensis the basal membrane is present (commonly c. 87 μm high) and the segments are well-developed (from c. ¾ to nearly equal the height of the teeth and narrowly perforate). Cilia are absent in both species. Stomata in E. trentepohlioides are exceedingly difficult to observe, cryptopore, and either few and scattered or none at the capsule base, while those of E. tjibodensis are relatively easily observed, phaneropore, numerous, and arranged in a single transverse row. In E. trentepohlioides the spores are mostly transversely septate, whereas those of E. tjibodensis are unicellular. Ephemeropsis trentepohlioides has a more compact protonemal weft than E. tjibodensis. The protonema of E. trentepohlioides lacks both the holdfasts ("Hapteren") described by Renner (1935) and Fleischer (1908) and the elongate terminal protonemal filaments ("Assimilatoren") described by Renner (1935) that are characteristic of E. tjibodensis. Ephemeropsis trentepohlioides is autoicous (with ♂ and ♀ gametangial buds connected by protonemal filaments), while my observations tend to confirm Fleischer’s (1908) statement that E. tjibodensis is either dioicous or occasionally apparently monoicous.
| Category | Number |
|---|---|
| Indigenous (Non-endemic) | 1 |
| Total | 1 |