Class
Family
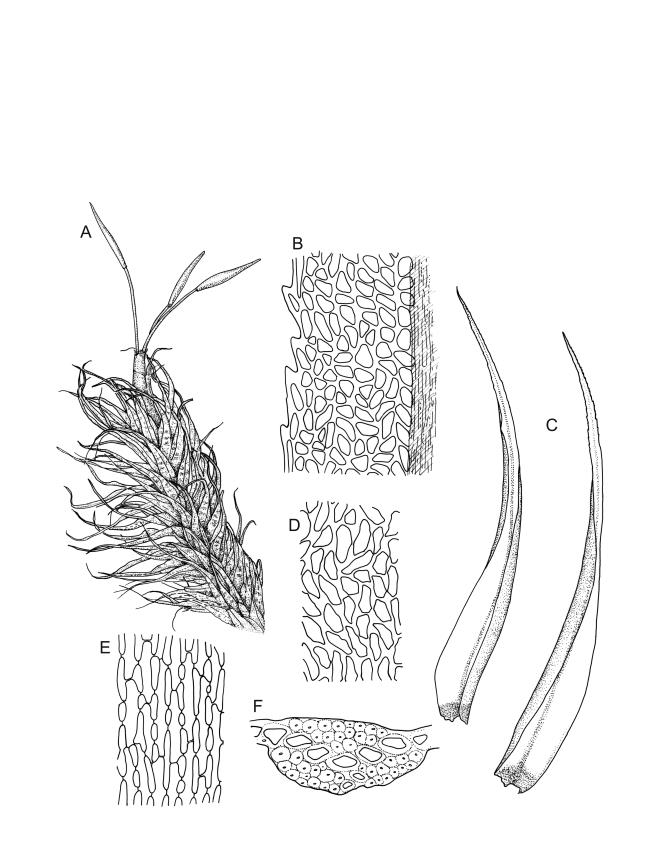
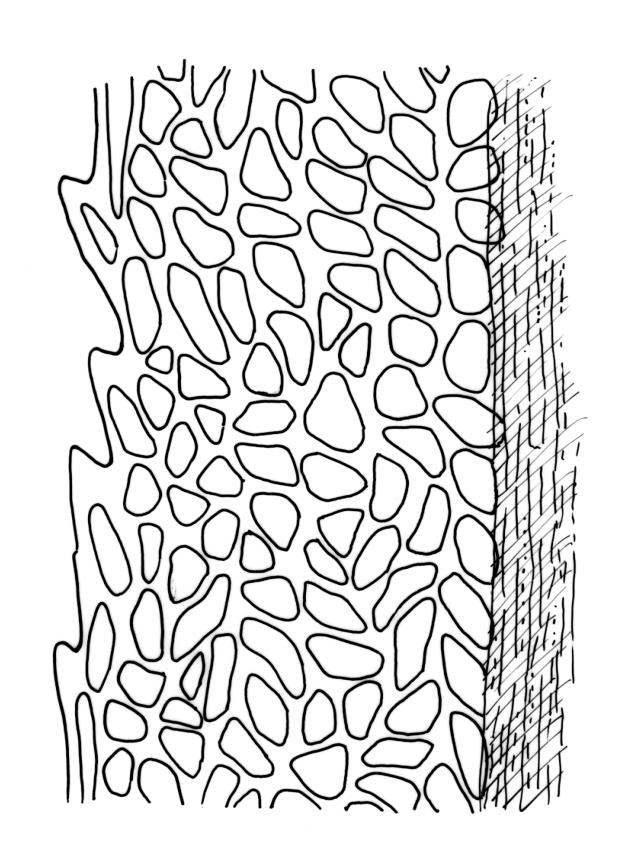
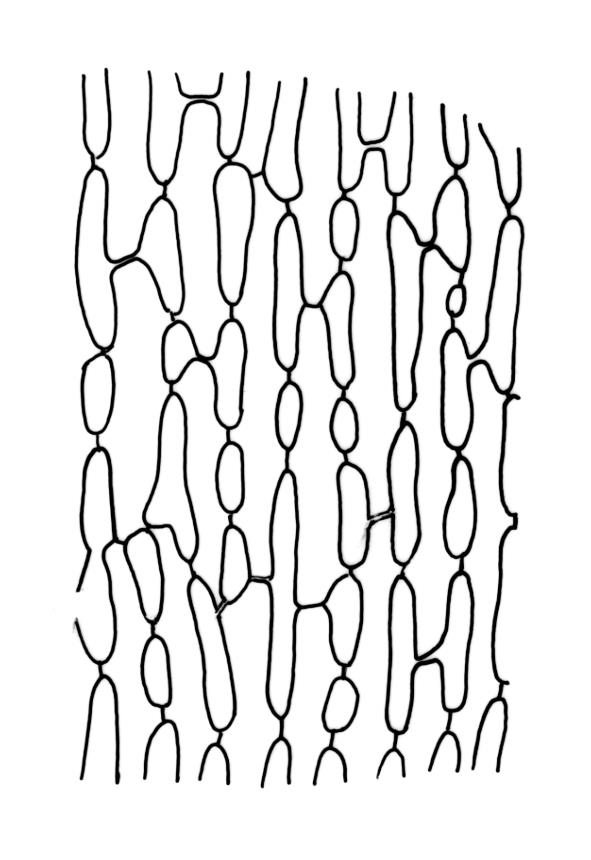
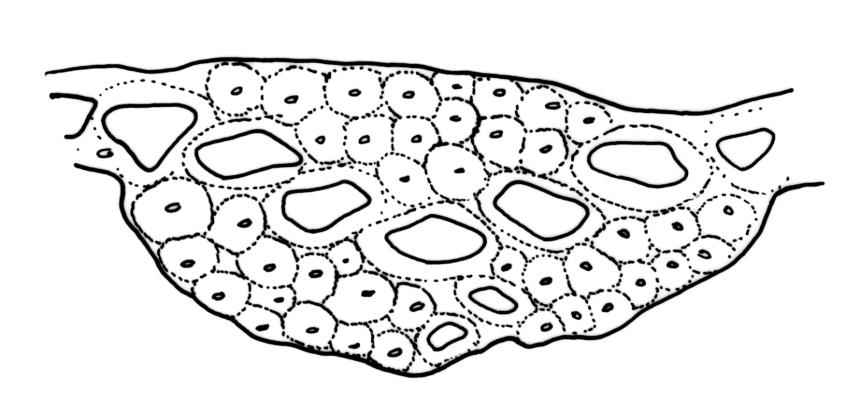
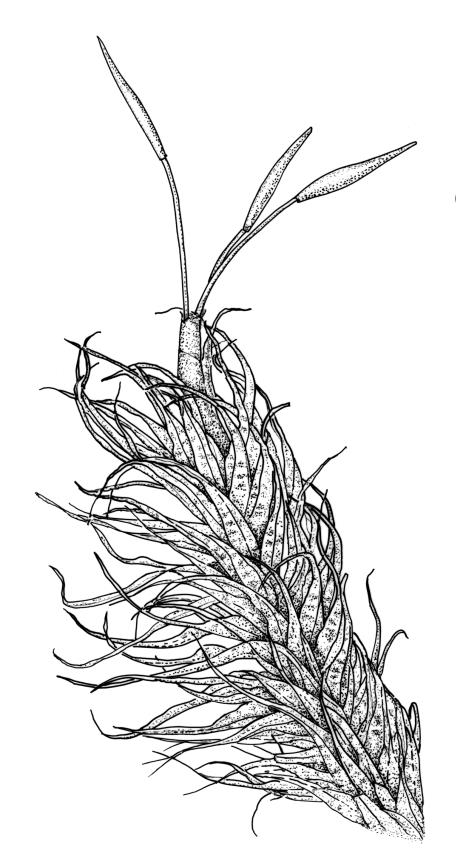
Plants robust, white-green when fresh, becoming gold-brown in older dried specimens, forming cushions or turves. Stems c. 35–75 mm, mostly sparsely forked, in cross-section with several layers of poorly differentiated but firm-walled cortical cells and a weak central strand, beset throughout by short, tangled, smooth, pale brown rhizoids. Leaves secund, weakly or strongly falcate, rugose, not plicate, becoming flatter and appearing wider but otherwise little altered when dry, ovate-lanceolate and ± narrowed to a clasping base, toothed (but not spinose) in upper ⅓ or less, mostly 8–9(–11) × 1.4–1.5 mm (under cover slip), subtubulose, plane at margins; mid laminal cells highly irregular in shape (a mixture of oblong, rhombic, triangular, oval, and irregular) and arrangement, mostly < c. 30 µm long and c. 12 µm wide, thick-walled and moderately porose, with short and irregular cells extending to apex; juxtacostal cells at mid leaf not differentiated; border well defined and extending from just above alar group to base of teeth, c. 8 cells and 36–42(–60) µm wide at mid leaf, gradually narrowing above; cells of leaf base elongate, mostly 45–90 µm, extremely porose; alar cells strongly and abruptly differentiated, unistratose, extending c. ¾ or more to costal base and c. 10 cells up the margin, inflated and yellow, firm-walled, the largest c. 90 × 40 µm. Costa narrow, occupying c. 1/15 of widest part of leaf base and c. 100 µm wide, mostly excurrent, the abaxial surface in upper ¼ or less bearing 2 rows of weak spines (not obvious under hand-lens), in cross-section lacking wings at mid leaf, with a median layer of c. 6 guide cells and one abaxial and one adaxial stereid group, which extend the width of the costa.
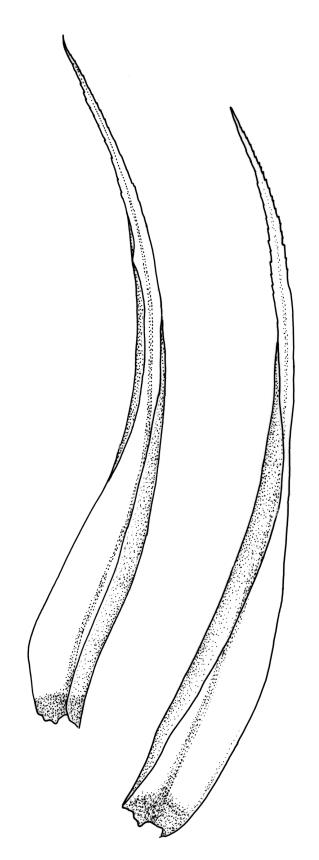
Pseudautoicous. Perichaetial leaves strongly sheathing (c. ½ the seta length) and tubulose, elliptic, rapidly tapered to a rounded apex, with the costa narrow and long-excurrent to form a slender arista c. ⅓ to ½ the length of the base. Dwarf males gemmiform, embedded in rhizoids of sterile or ♀ plants. Setae aggregated, (1–)2–4 per perichaetium, 10–11 mm, straight, not twisted, pale or red-brown; capsules exserted, curved, cylindric, 2.9–3.4 mm, weakly strumose, smooth when moist, constricted below the mouth and weakly sulcate when dry; exothecial cells firm-walled and oblong-hexagonal; stomata restricted to neck, superficial; annulus weakly differentiated, not revoluble; operculum curved-rostrate from a conic base, ± equal to the capsule. Peristome teeth inserted at mouth, red-brown, split c. ½ to base into segments of unequal width (often with 1–2 elongate perforations at the base of the division), c. 525–640 × 100 µm, papillose-striolate in lower ½ to ⅔, baculate near apex. Calyptra cucullate, smooth. Spores spherical, firm-walled, 18–22 µm, nearly smooth.
Dixon 1913, pl. 2, figs. 10a–d; Tan 1989, figs. 15–18; Malcolm & Malcolm 2006, pp. 103, 107, 158, 162, 171, 244, 247.
Dicranoloma platycaulon is a relatively uncommonly collected species. It is most often confused with D. plurisetum, which has a similar pale white-green colour when fresh. The leaf undulations, the decidedly less strongly toothed upper margins, and the irregularly shaped and oriented upper laminal cells in the present species are sufficient to preclude confusion. The relative flatness of the leaf base in dry material of D. platycaulon also aids in its separation from D. plurisetum. According to J.E. Beever (pers. comm., 1 Aug. 2017) D. platycaulon has red, immature setae, while those of D. plurisetum are green or yellow.
NI: N Auckland (Tūtāmoe Range), S Auckland (Moehau, Pirongia), Gisborne (Mt Maungapōhatu, Lake Waikaremoana, Manuoha Ridge), Taranaki (Mt Taranaki), Wellington; SI: Nelson, Marlborough (Kenepuru Valley), Canterbury (Arthur’s Pass), Westland, Otago, Southland; St.
Australasian. Tasmania*, mainland Australia (Victoria*). Reported from Borneo and New Guinea by Tan (1989), but these reports are doubted by Klazenga (1999, pp. 57 & 122, and, by omission, Klazenga 2003). However, as suggested by Tan, the occurrence of a predominantly Australasian species at high elevations in Malesia seems phytogeographically plausible.
On duff or rock (granite or greywacke) on forest floor and on tree roots and bases. Usually forming cushions (to c. 30 cm diam.) on soil or rock hummocks, but also forming relatively small turves. Occurring mostly in forest with one or more species of southern beech. Also occurring as an epiphyte on trunks of Fuscospora solandri s.l., Lophozonia menziesii, Weinmannia racemosa, and the tree fern Dicksonia squarrosa. On the North I. no low elevation occurrences have been documented but ranging to 1220 m (Manuoha Ridge) and on the South I. from near sea level (Punakāiki River and Lake Moeraki, both Westland L.D.) to 1350 m (Glasgow Range, Nelson L.D.). Occurrences below c. 500 m elevation are primarily epiphytic. Frequently associated with Dicranoloma plurisetum, D. robustum, Leptotheca gaudichaudii, as well as Bazzania adnexa, and Lepidozia spp.
In the field the pale coloration and the distinctly rugose aspect of the leaves are distinctive. Microscopically, the laminal cells, from c. mid leaf to the apex, are highly irregular in form, and the orientation of one cell usually bears little relationship to that of adjacent cells. The resultant confused jumble of cells gives the lamina a highly distinctive microscopic appearance unlike any other N.Z. Dicranoloma. The distance to which the randomly oriented and highly irregular cells extend towards the leaf base varies considerably between populations. In the type, the highly irregular cells of the upper lamina extend downward to mid leaf or lower. In much material the laminal cells are in alternate, vaguely defined transverse bands of randomly oriented irregular cells and more uniformly oriented and more uniformly shaped cells. The cell bands appear to be related to the characteristic leaf undulations. The leaves in this species become less concave (and hence appear broader) at the base when dry.
Epiphytic forms from lower elevations in Nelson L.D. (e.g., A.J. Fife 6041 from Punakāiki River, CHR 405710) have larger (c. 11 mm × ≥2.0 mm wide) and more setaceous leaves, with cells in mid lamina and above longer (mostly 27–42 µm) and more porose than in type material. The upper leaves are ± more sharply toothed than in terrestrial populations, and the leaves are often brittle and fragile. However, in these forms the leaves are strongly undulate and have bands of highly irregular laminal cells interspersed in the upper half or more of the leaf. Such large-leaved epiphytic forms are best referred to D. platycaulon and can be considered analogous to the so-called "setosum" growth form of D. robustum.
Considerable confusion surrounds the publication and typification of the name D. platycaulon. The epithet platycaulon was first applied, in herb., by Müller Hal. to R. Helms 71 from Westland. Beckett (1899) published "Dicranum platycaulon" but his "protologue" combined Müller’s name with a diagnosis written by Brotherus using another specimen (T.W.N. Beckett 494 from Patterson’s Creek, Canterbury L.D., CHR 585951). Wijk et al. (1962, p. 91) interpreted "Dicranum platycaulon Müll.Hal. in Beckett" as a nom. nud.; their interpretation is accepted here. A portion of R. Helms 71 is present in the Beckett herbarium (CHR 585951), and it is clearly referable to D. plurisetum Dixon. Two of the other six syntypes (leg. Jupp, Bloxam) cited by Dixon are correctly referable to D. platycaulon.
Dixon (1913, p. 15) provided the first valid publication of D. platycaulon. Dixon’s reference to Müller’s nom. nud. as a basionym is both superfluous and confusing. Dicranoloma platycaulon should be attributed to Dixon alone, as done by both Wijk et al. (1962, p. 47) and Klazenga (2003). However, Dixon’s designation of a type is confusing. He cited a "type in herb. C. Müll., in Mus. Bot. Berolin." but in a footnote stated that he had not seen "the Greymouth specimen" (a reference to R. Helms 71) and that his "description and figures are drawn up from Bell’s Mount Cargill plant, sent me by Dr Brotherus." W. Bell routinely sent his Otago moss collections to Beckett, who then forwarded them to other bryologists, including Müller and Brotherus. Tan’s (1989) designation of the Mt Cargill material as the lectotype is accepted here. His illustration of the Bell collection leaves no doubt as to the taxonomic identity of the material he examined, and the lectotypification preserves current usage.
An apparent isolectotype is CHR 543119, as herb. Beckett 640. Although the specimen is labelled in Beckett’s hand as collected by D. Petrie, it also bears a handwritten citation "Mt. Cargill" in W. Bell’s hand, suggesting that Bell was probably the actual collector. The specimen is ample and fruiting.