Class
Family
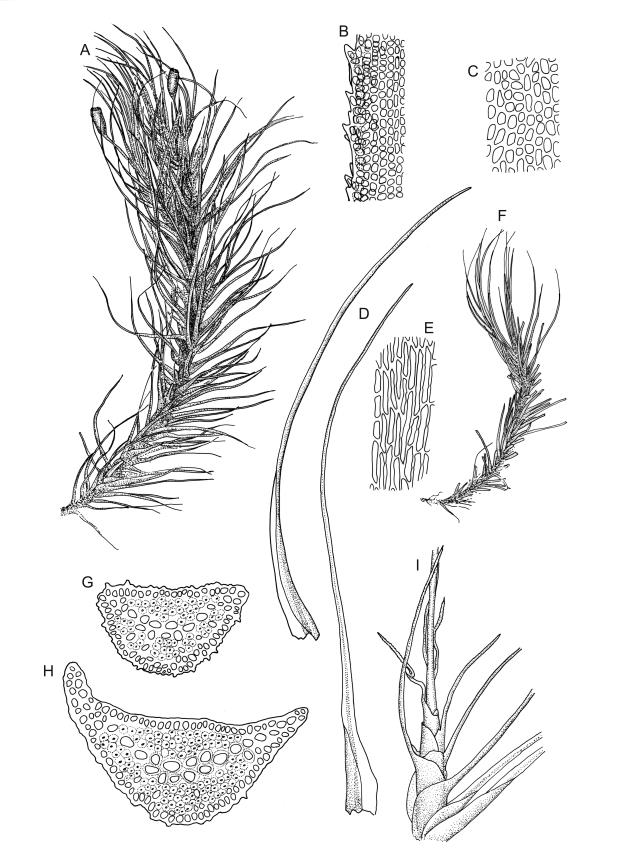
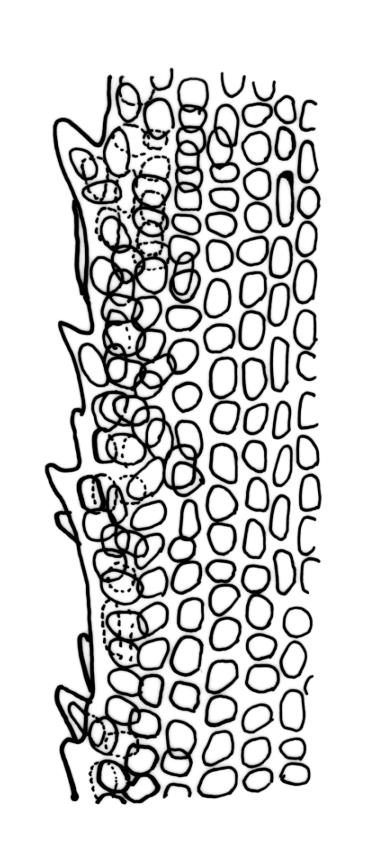
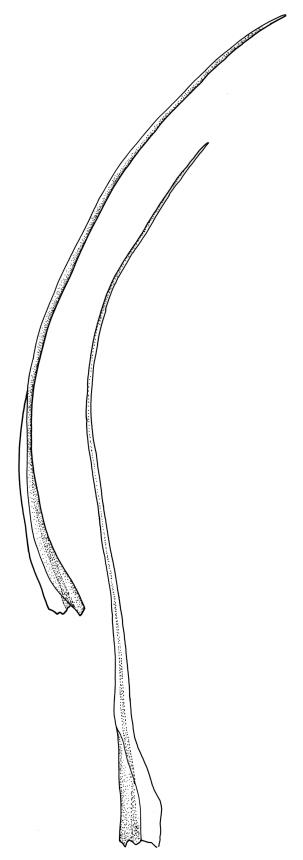
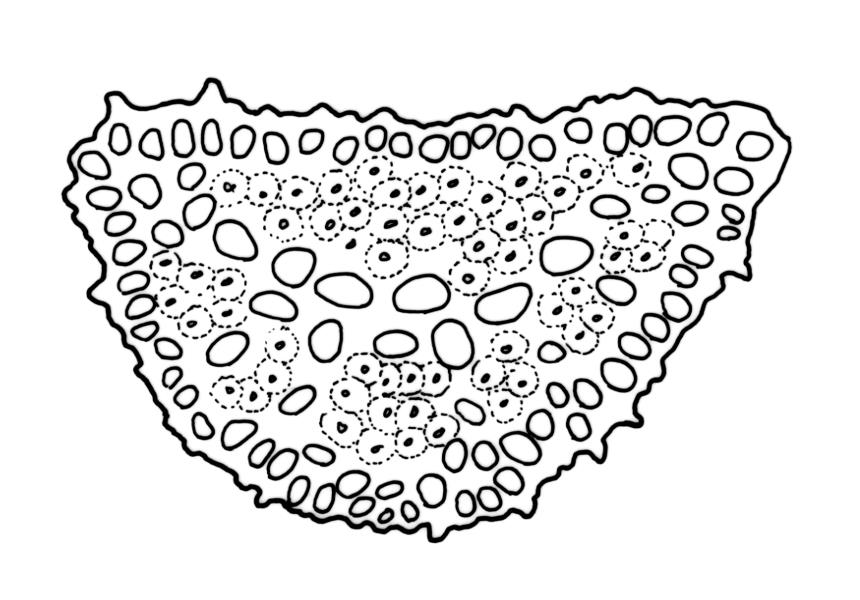
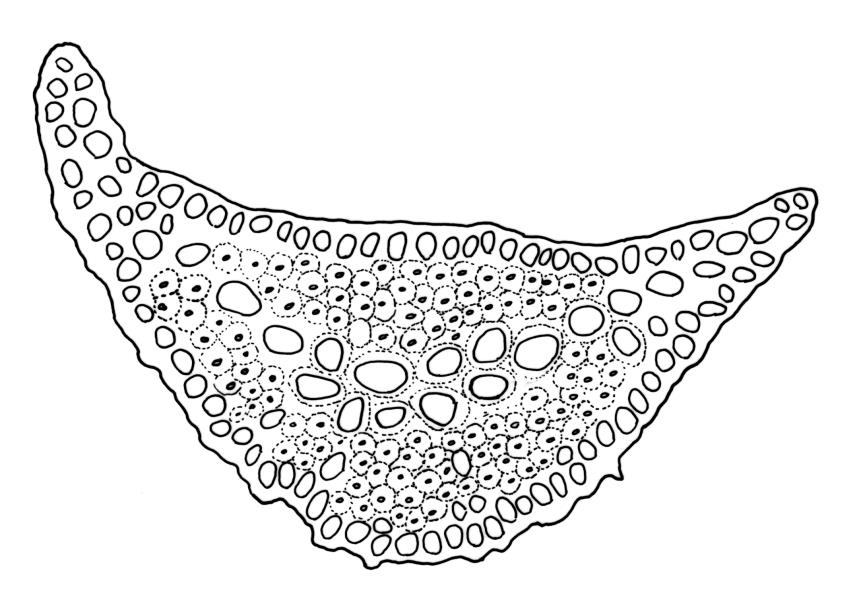
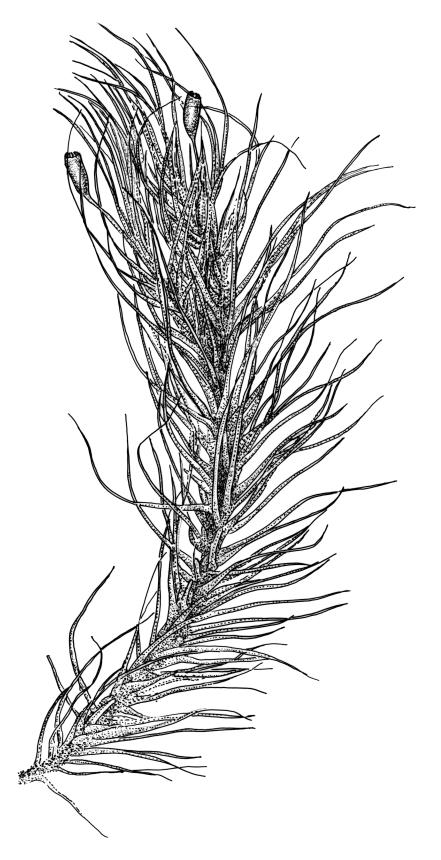
Plants robust, dark green or yellow-green and dull when fresh, becoming gold-brown in older dried specimens, forming loose cushions or extensive turves. Stems (10–)35–50 mm, branching by subperichaetial innovation and by forking, in cross-section with a weak and ill-defined central strand and several ill-defined layers of cortical cells; matted throughout by short, tangled, smooth, and pale brown to nearly white rhizoids. Leaves mostly weakly secund, sometimes rigid and erect, occasionally falcate, neither plicate nor rugose, linear-lanceolate from a short ± oblong base, to a long-setaceous apex, which is ± filled by the costa and often fragile, sparsely and weakly serrate in upper ½ or less, 9–11(–14 or greater) × 0.7–0.9(–1.2) mm (under cover slip), subtubulose below, carinate above, plane at margins; mid laminal cells isodiametric, short-rectangular or short-oblong, mostly 8–12 × 6–8 µm and 1:1–2, thick-walled, not porose; juxtacostal cells at mid leaf often bistratose but not otherwise differentiated; border ill-defined, consisting of 1–3 weakly differentiated rows of ± elongate cells or sometimes lacking; cells of leaf base mostly elongate (30–55 µm) but sometimes shorter, usually porose; alar cells strongly and abruptly differentiated, unistratose, extending to costal base and c. 6–12 cells up the margin, inflated but firm-walled, mostly rectangular, yellow-brown or hyaline. Costa broad, filling c. ⅕–⅓ the widest part of leaf base, the abaxial surface with cell ends weakly projecting (to c. mid leaf), the resultant weak spines not visible under the hand-lens, in cross-section lacking wings at upper end of leaf base, c. 8–10 cell layers thick, with a median layer (–2 layers) of c. 10–12 guide cells, well-developed abaxial and adaxial stereid groups, which extend the width of the costa, and a single layer of slightly larger-lumened cells on both surfaces (sometimes with 1–3 isolated larger cells amongst the abaxial stereid group).
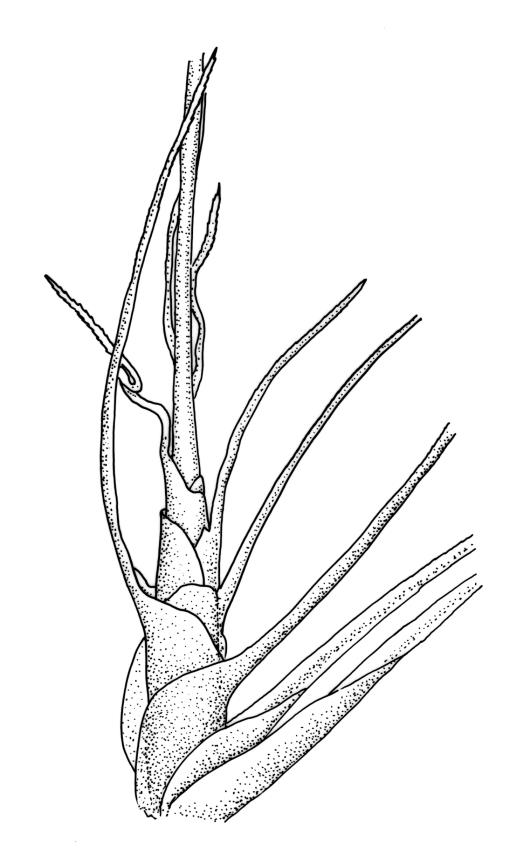
Pseudautoicous (♂ plants occasionally not dwarfed). Perichaetial leaves sheathing and strongly concave, abruptly tapered from an elliptic base to an obtuse or rounded apex, with costa present in base and excurrent to form a long and slender arista that is usually 1–2 times the length of the base (the outer ♀ leaves often with aristae up to 4 times the base length). Dwarf ♂ plants embedded in rhizoids of sterile or ♀ plants; non-dwarf males occasionally seen. Setae 1 per perichaetium, 4.5–6(–10) mm, straight, not twisted; capsules strongly curved, ± cylindric, 2–2.5 mm, not or weakly strumose, ± smooth when moist, constricted below the mouth and sulcate when dry; exothecial cells firm-walled and oblong-hexagonal; stomata restricted to neck, superficial; annulus not seen; operculum curved-rostrate from a conic base, ± equal to the capsule. Peristome teeth inserted at mouth, red-brown, split c. ½ to base into segments of unequal width, c. 465–525 × 100 µm, longitudinally striolate in lower ½ or more, baculate near apex. Calyptra cucullate, smooth. Spores spherical, 15–18 µm, nearly smooth.
Wilson & Hooker 1845, pl. 58, IV (as Dicranum menziesii); Klazenga 2003, fig. 19; Meagher & Fuhrer 2003, p. 143.
Dicranoloma menziesii, in particular the "rigidum" growth form, is likely confused with the "setosum" growth form of D. robustum. However, in all instances the consistently shortly rectangular (<2:1) or quadrate subula cells of D. menziesii differentiate it from D. robustum "setosum" growth form, in which the laminal cells in the mid portion of the subula are seldom less than 30 µm long, nor less than 4-5:1. The non-porose nature of the mid laminal cells in D. menziesii also contrasts with the porose cells in the D. robustum "setosum" growth form. The difference in laminal cell shape can generally be seen under the stereoscope, but microscopic examination can be necessary for certain identification.
NI: N Auckland, including offshore islands (PK, HC, LB, GB), S Auckland, Gisborne, Hawke’s Bay (Puketītiri), Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; Sol; Sn; A; C.
Austral. Tasmania*, mainland Australia*. Recorded from Lord Howe I., southern South America, and New Caledonia by Klazenga (2003).
Exposed roots, tree trunks, stumps and logs, rock, occasionally on soil or duff (but not forming extensive turves on duff). Occurring on trunks of a wide variety of coniferous and broad-leaved trees, shrubs, and tree ferns in virtually all kinds of forest types from near sea level to the subalpine. As an epiphyte forming cushions up to at least 40 cm diameter. From near sea level (Tī Point, N Auckland L.D.) to at least 1160 m (Mt Pihanga near Turangi, Wellington L.D.) on the North I. and from near sea level (several localities) to at least 1210 m (Waterfall Creek near Maruia Springs, Nelson L.D.) on the South I.
Dicranoloma menziesii is exceedingly widespread and is common in most forest types; it is highly variable, particularly with respect to the length and orientation of its leaves, the relative fragility of the leaves, and the coloration of fresh plants. The most consistently useful field recognition feature is the short sporophyte, which is exceeded in length by both the vegetative and outer perichaetial leaves. The inner perichaetial leaves are much shorter and sheathe the setae to variable heights. The very narrow leaves with broad and thick costae and fine setaceous apices are generally sufficient to distinguish it from its congeners, with the exception of the "setosum" growth form of D. robustum. D. menziesii is the only N.Z. species of its genus with ± isodiametric mid to upper laminal cells.
Non-dwarf male plants, equal in stature to associated female plants, occur in this species (e.g., D. Petrie s.n., 20 Nov. 1896 from Waingaro, S Auckland L.D., CHR 543118). The perigonia here are terminal on stems and branches, have bracts much reduced relative to the vegetative leaves, and enclose numerous filiform paraphyses. Ramsay (1985), in an investigation of the cytological and physiological factors involved in sexual differentiation in Dicranoloma, also reported non-dwarf male plants in this species.
No type material of Dicranum menziesii Taylor from Norfolk I. has been seen. Taylor’s publication of this name precedes the previously accepted name D. menziesii Hook.f. & Wilson by one month according to Klazenga (2003), whose dates and consequent nomenclature are accepted here. The two isotypes of Dicranum menziesii Hook.f. & Wilson seen are both sterile and consist of single stems with leaves very weakly secund and 7–10 mm.
Two probable isotypes of Dicranum menziesii var. rigidum have been examined. One is filed as Dicranum fasciatum in NY-Mitten and is labelled "Dusky Bay, New Zealand, leg. & comm. A. Menzies" in what is apparently Menzies’s script. That in BM is filed under Dicranoloma menziesii, lacks varietal designation, bears a "type" stamp, and is labelled simply "New Zealand, Menzies". Both these specimens have sporophytes and very fine, non-fragile, and slightly secund leaves of 8–9 mm.
Some synonyms of Dicranoloma menziesii are included in this treatment based on work of earlier authors. Dicranum trichophyllum Hampe and Dicranum fulvum R.Br.bis, hom. illeg. were synonymised by Dixon (1913)and subsequently confirmed, in one case tentatively, by Klazenga (2003); they are accepted here.
Dicranodontium australe Dixon [Bull. New Zealand Inst. 3(3): 92, 1923] was initially synonymised with Dicranoloma menziesii by Sainsbury (1946). Klazenga (2003) proposed a lectotype from Great Barrier I. A probable isolectotype (CHR 542644!) has been studied and Sainsbury’s synonymy is affirmed here on this basis.
Dicranum calymperoideum Müll.Hal. [Hedwigia 36: 359, 1897] was proposed as a synonym by Robinson (1975, p. 22). It is based on a Beccari collection from the Titirangi Range near Auckland and was not confirmed by Klazenga (2003). It is excluded from the list of synonyms here.
Populations in which the leaves are rigidly erect and sometimes fragile have been termed var. rigidum. In some populations the erect leaves can exceed the length cited in the description, reaching c. 22 mm. Due to the very high number of morphological intermediates between rigid-leaved populations and those with loosely secund leaves, the var. rigidum is considered here to be a growth form of D. menziesii; this accords with the taxonomy of Klazenga (2003).
Dicranoloma menziesii occasionally forms detached "moss balls". J.S. Thomson (in litt. to L.B. Moore, 18 June 1941, filed with CHR 263240) described seeing "some 50 specimens" from Stewart I. Most of the unattached "moss balls" occurred in a "more or less open space about 12 ft. in diameter on the floor of [forest]… at the head of Paterson’s Inlet. The largest specimen observed measured about 8" in diam., but the majority were about 4", 5", or 6" in diameter." Similar "moss balls" of D. menziesii have been found in the Kakeahua Valley on the same island by R. Buckingham (cf. CHR 242724). R. Buckingham suggested (pers. comm.) that they may be caused by "disturbance associated with feeding or display activities of birds".