Class
Family
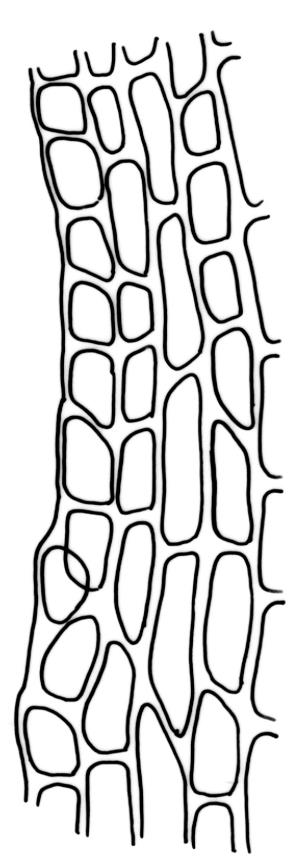
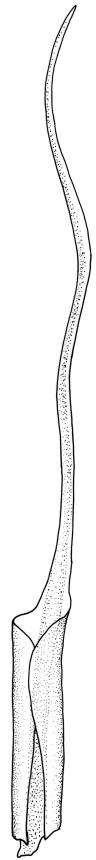
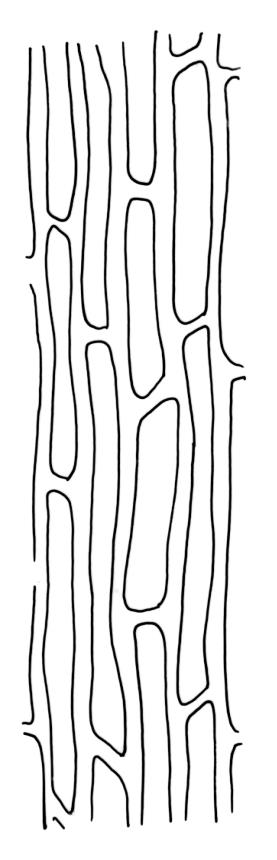
Plants bright green, often becoming brown in older herbarium material. Stems variable in length, commonly c. 2–6 mm, but sometimes to c. 40 mm. Leaves sheathing at base and shouldered. Lower leaves mostly c. 2.0–2.5 mm, sheathing at base, tapered abruptly from an oblong base to a narrow subula that is ⅔ or more the total leaf length, acuminate at apex, entire or finely denticulate at apex, channelled below, unistratose. Upper (including perichaetial) leaves c. 3–6.5 mm, strongly sheathing, in shape as the lower leaves, the subula c. ⅔ the total leaf length and nearly filled by the costae above (lamina at mid subula c. 4 cells wide). Upper laminal cells (in leaves from below perichaetium, mid leaf) absent or to 6 cells wide, 12–18(–21) × c. 6 µm, and unistratose at margins. Costa (in leaves from below perichaetium) occupying c. ⅙ the width and ± poorly defined in leaf base, sometimes appearing to fill the upper subula.
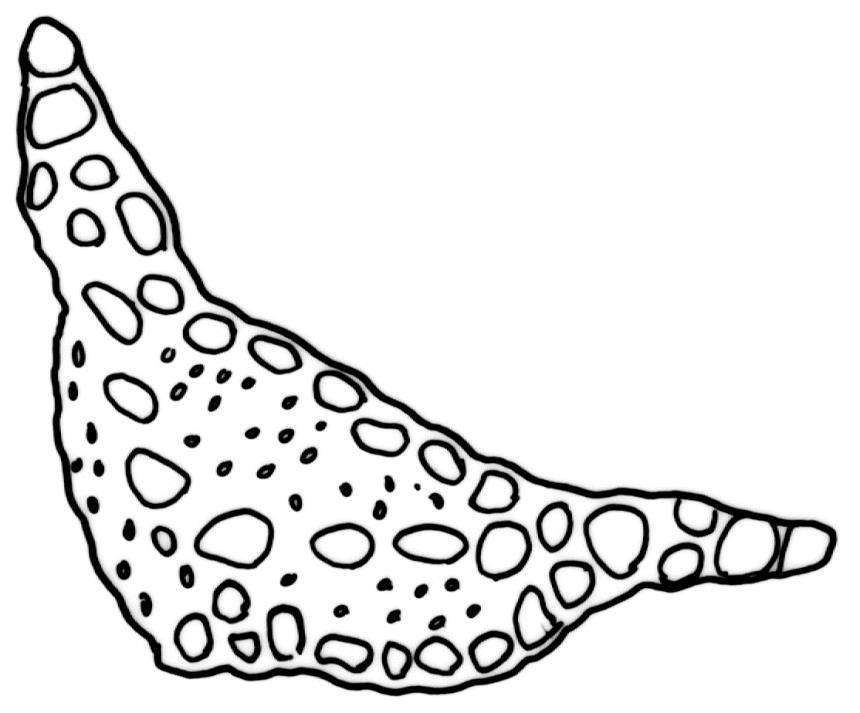
Dioicous. Perichaetia often multiple and sequentially overtopped by innovations, with leaves as above. Perigonia often inconspicuous. Setae c. 2.5–4(–6) mm, yellow-brown; capsules emergent (not or scarcely exceeding perichaetial leaves), ovoid to short-cylindric, 0.8–1.2(–1.5) mm, slightly asymmetric at base, with a variably developed sterile neck and usually weakly strumose; exothecial cells and annulus as per genus; stomata superficial, restricted to capsule base; operculum c. 0.5 mm. Peristome as per genus. Spores 22–29 µm.
Giese & Frahm 1986, figs. 1–22; 37–41.
Confusion sometimes occurs between C. capillaceum and the relatively rare Dicranella dietrichiae, with sterile material being especially confusing. Campylopodium capillaceum has longer (c. 3–6.5 mm vs c. 4 mm) and more slender upper and perichaetial leaves that are not contorted when dry compared to the shorter and much-contorted dry leaves of D. dietrichiae. When fertile, the shorter, stouter, and distinctly cygneous setae of C. capillaceum preclude confusion. Finally, stomata in C. capillaceum are conspicuous and phaneropore, while in D. dietrichiae stomata appear to be absent.
Confusion with Dicranella cardotii is less likely, with differences in plant stature, colour, and leaf shape helping to distinguish C. capillaceum. Microscopically the mid leaf laminal cells of C. capillaceum are unistratose and smooth, compared to the mostly bistratose and bulging-mammillose cells of D. cardotii. Dicranella cardotii is also largely confined to stream margins and seepages, at least in N.Z.
Some collections of small forms of Campylopus pyriformis named as C. capillaceum/medium have been seen. Campylopodium capillaceum medium has narrower costae with adaxial stereids, shorter and stouter setae, larger spores, non-fringed calyptrae, and conspicuous stomata in the capsule neck. Stomata are absent in Campylopus pyriformis, as they are in all members of that genus.
NI: N Auckland including offshore islands (LB, GB), S Auckland, Gisborne (Panikau), Hawke’s Bay (Wairoa), Taranaki (Tāngarākau Gorge), Wellington; SI: Nelson, Canterbury (Arthur’s Pass), Westland, Otago (several localities near Dunedin), Southland; St.
Anomalous. Tasmania*, Java*, Samoa*, Tahiti*, Austral Is.*. Also reported from Sumatra, Celebes, Malaysia, Philippines, New Guinea, Fiji, Hawai‘i, New Caledonia, Taiwan, Japan, Chile, and Puerto Rico by Giese & Frahm (1986).
On mineral soil, especially clay, in a wide range of vegetation types. Very often on vertical sides of drainage ditches and track- or road-side banks, but also on ± bare soil patches. Occasionally on thermally heated soil, on soil at lake or stream margins, and in pākihi. Apparently avoiding calcareous (or nutrient enriched?) sites. On the North I. from near sea level (Okura Bush, N Auckland L.D.) to at least 1036 m (Table Top in Tararua Range, Wellington L.D.) and on South I. from near sea level (Punakāiki area, Nelson L.D.) to 1400 m (Scarlett Range, Nelson L.D.).
Although most often a small plant with stems <6 mm, C. capillaceum is extremely variable, particularly in regard to stature and the capsule shape and dimension. Capsule length can vary considerably, even in single populations. The extent of the sterile neck tissue and the associated weakly developed struma at the capsule base is also highly variable. Forms with elongate stems (20–40 mm or greater) are usually from thermally heated sites or subject to seepage. In these conditions, stems sometimes proliferate repeatedly by subperichaetial or subperigonial innovations.
Like Sainsbury (1955, p. 98), I have not seen the type of Campylopodium euphorocladum var. thermophilum Dixon & Thér. in Dixon & Sainsbury [J. Bot. 71: 215, 1933] but have studied the same Allison collection (K.W. Allison 608, CHR 543122A) from Rainbow Mountain that Sainsbury cited. It falls within the continuous range of variability for the species in all features except stem length, and I do not consider it worthy of taxonomic recognition. The type collection of the variety was collected at "Rainbow Hill" in Sept. 1929 by A. Meebold.
The name Campylopus capillaceus Hook.f. & Wilson is an earlier valid name for what has been termed Campylopodium medium (Duby) Giese & Frahm (Giese & Frahm 1986). A packet of type material of Campylopus capillaceus Hook.f. & Wilson, containing several fruiting plants, is present in herb. Wilson (as "W. 322"). It is annotated by Wilson as "Campylopus capillaceus Mitt." and accompanied by Wilson pencil sketches. It is attached with several other duplicates of the collection J.D. Hooker 322 (from Bay of Islands) to BM sheet 000079658.
In Flora Novae-Zelandiae (Wilson 1854, p. 67) Hooker material from the Bay of Islands is referred to, under the genus Dicranodontium, as "D. proscriptum Hornsch. var. β". The allegedly earlier name mentioned by Wilson, Cynodon proscriptus Hornsch. [Horae Phys. Berol. 60: 357, 1820], was based on material from St Helena. It was excluded from Campylopodium by Giese & Frahm (1986, p. 133) and is not considered further here.
Both Campylopodium medium (Duby) Giese & J.-P.Frahm and C. euphorocladum (Müll.Hal.) Besch. appear to be based on segregates of a single Javanese collection made by H. Zollinger (see Giese & Frahm 1986).
Giese & Frahm (1986, p. 128) discussed Japanese material of Campylopodium capillaceum (as C. medium) in which the peristome teeth are "either entire or the forks are imperfectly divided." Material from Mt Owen (J.K. Bartlett 22102, CHR 360613) is rather poor material of C. medium, having peristome teeth appearing undivided and coarsely striate throughout. All other features of this sparse collection point convincingly to its referral to this common and widespread species, rather than a new species of Dicranella, as Bartlett proposed (in herb.).