Class
Family
Subfamily
Genus
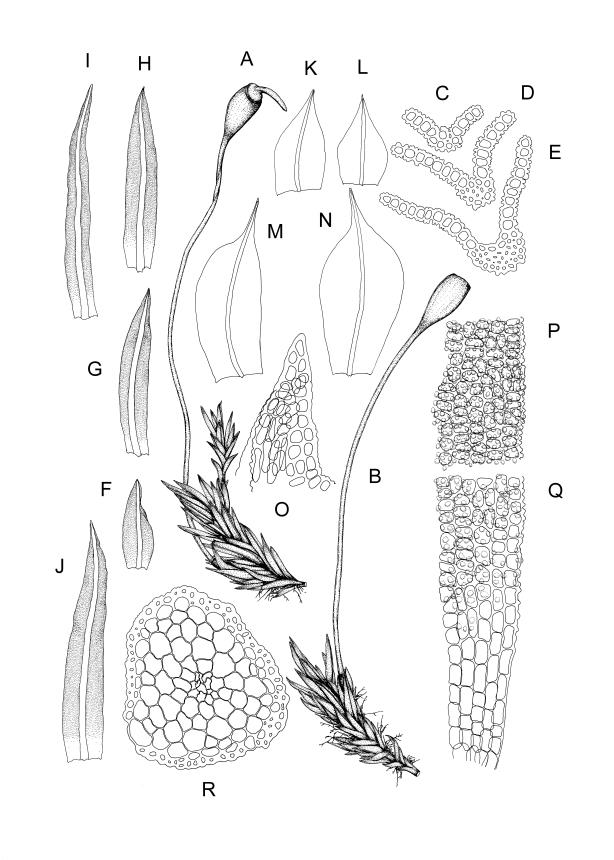
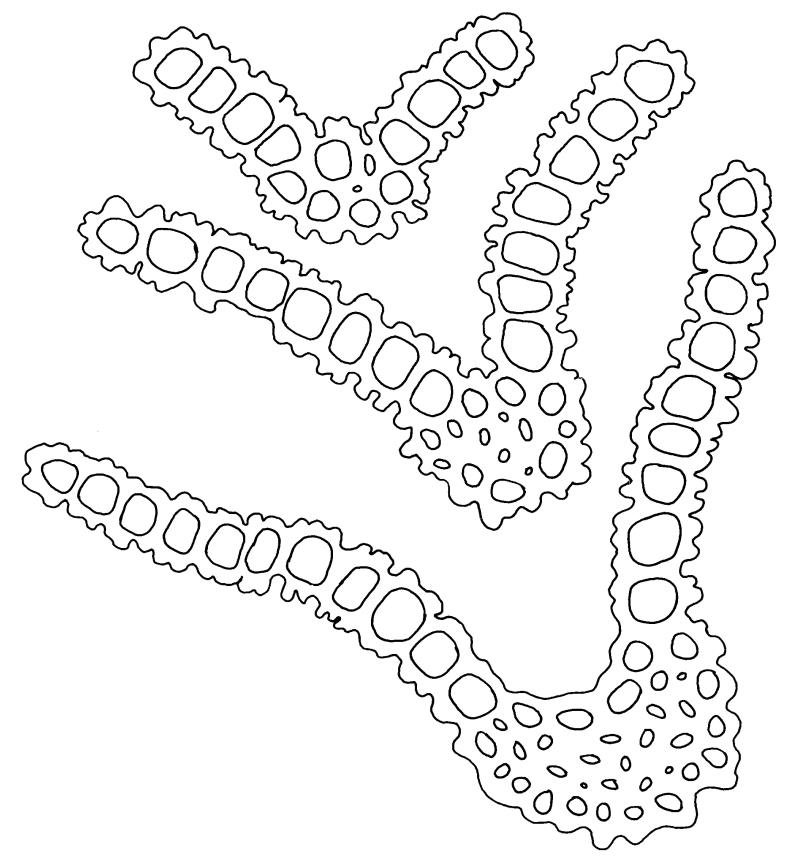
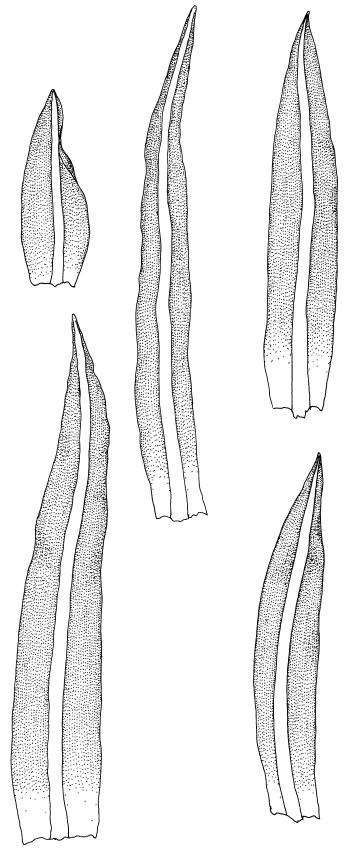
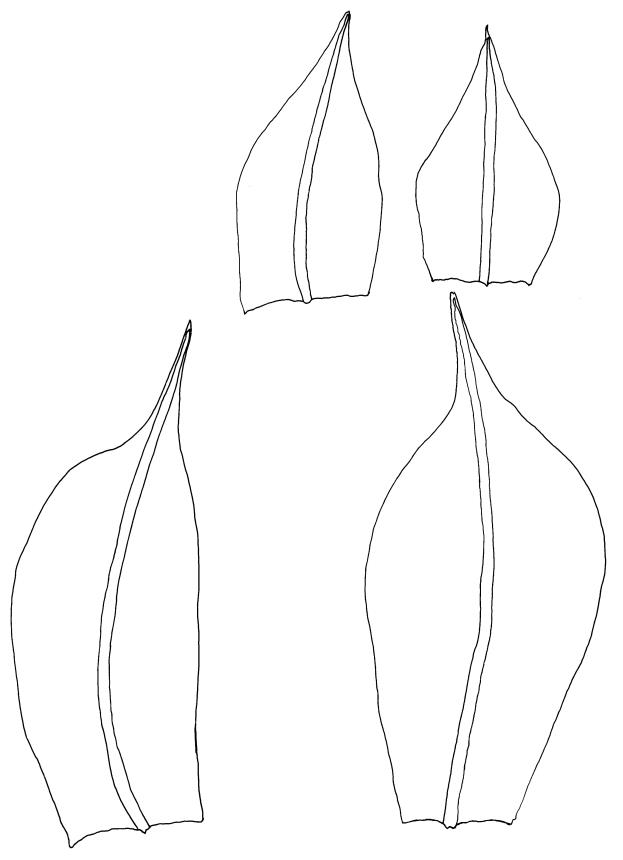
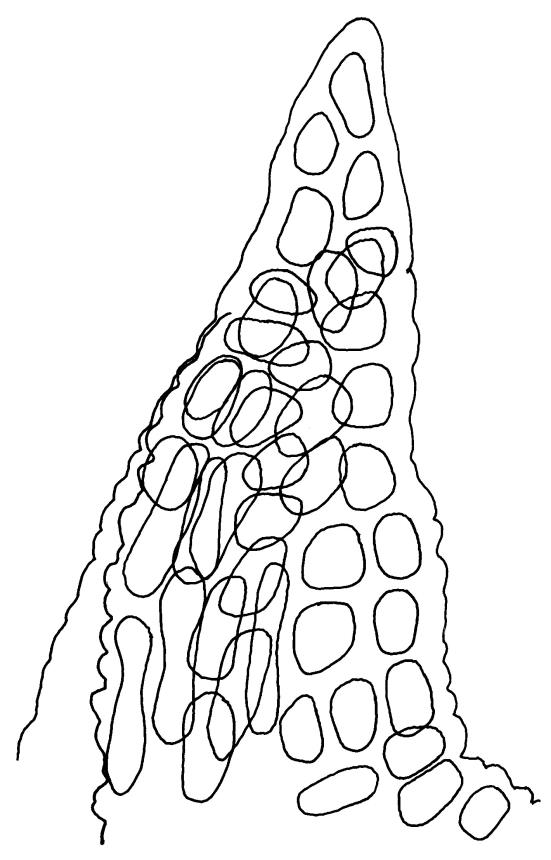
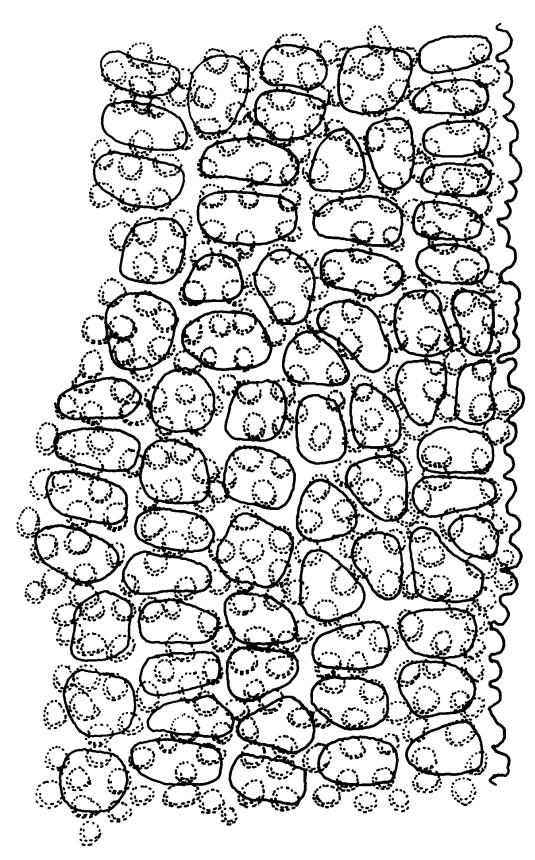
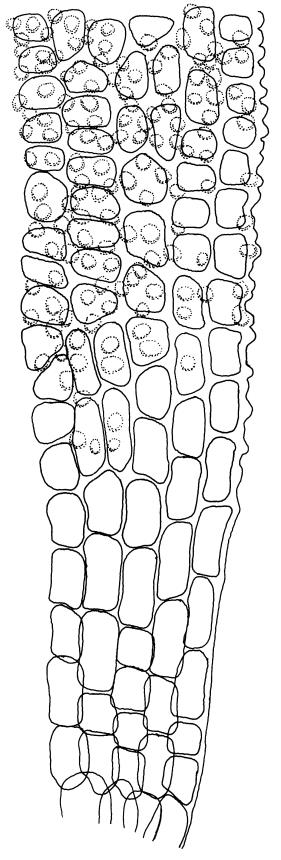
Plants bright yellow-green above, brown below, forming turves on damp rock. Stems 15–40 mm, simple or branched, usually matted together with a dense, red-brown tomentum, central strand present, sclerodermis well developed, hyalodermis absent. Leaves erect-spreading when moist, weakly individually twisted when dry, 1.0–2.5 mm, linear to lanceolate, carinate above, deeply grooved along the costa, acuminate; margins often weakly recurved below, papillose above; upper laminal cells obscure, subquadrate to elliptic, often oblate, thin- to thick-walled, densely pluripapillose with simple and complex papillae, (6.0–)9.0–12.0(–13.5) × (7.5–)9.0–10.5(–13.5) µm; upper marginal cells not differentiated; lower laminal cells differentiated only in extreme leaf base, quadrate to elongate, thick-walled, smooth. Costa concolorous, stout, percurrent to shortly excurrent, with 1–3 pellucid cells at the leaf apex, the terminal cell often bearing a single terminal papilla; adaxial superficial cells rectangular, papillose in distal ½, smooth in proximal ½; abaxial superficial cells fusiform, densely papillose throughout. Laminal KOH colour reaction yellow.
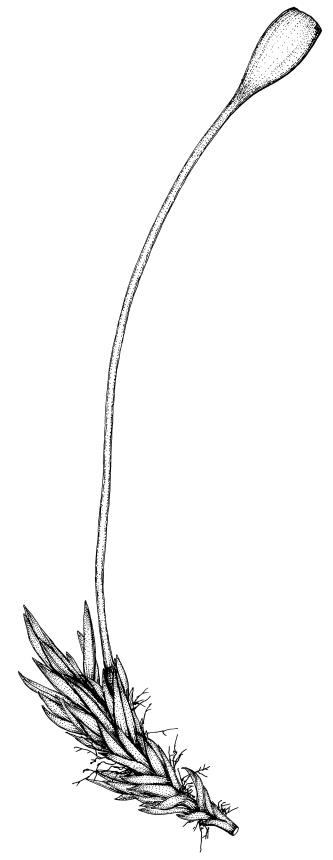
Reportedly dioicous (Zander et al. 2007). Perichaetia terminal on short lateral branches, with inner leaves subulate from broadly sheathing bases. Perigonia not seen in N.Z. material. Setae slender, 7–11 mm. Capsules erect, symmetric, ellipsoid, glossy, darker at the mouth, 1.0–1.2 mm. Operculum obliquely and narrowly rostrate, ⅔ to equal the theca length. Peristome lacking. Spores 13–17 mm, finely papillose.
Sainsbury 1955, pl. 25, fig. 1; Scott & Stone 1976, pl. 33 pro parte (as A. bellii); Crum & Anderson 1981, fig. 119, f–k; Zander 1993, pl. 38, figs 10–13; Lüth 2019, p. 504; Malcolm et al. 2020, p. 479.
Sterile material of Anoectangium aestivum has often been confused with Zygodon intermedius, but the latter species has the costa failing before the leaf apex, with papillae on the abaxial face of the costa confined to the distal portion (vs a percurrent to excurrent costa that is papillose abaxially almost throughout in A. aestivum). Sterile Amphidium cyathicarpum may also be problematic; in that moss there are numerous (c. 15–20) fine papillae on the upper laminal cells, and the cell outlines are not obscured (vs fewer, c. 6–10, robust, crowded papillae, that obscure the walls between the cells in Anoectangium aestivum). Amphidium cyathicarpum may also have irregularly denticulate leaf margins (vs margins entire, although papillose, in Anoectangium aestivum).
Confusion may also occur with Gymnostomum calcareum, but Anoectangium aestivum is a much larger plant, with longer leaves (1.0–2.5 mm vs <1.0 mm in G. calcareum). Leaf apices are acute to acuminate in A. aestivum, whereas in G. calcareum they are most often rounded-obtuse, but are variable in shape even within one shoot. The terminal cell is often unipapillose in A. aestivum (vs pluripapillose in G. calcareum).
Bryoerythrophyllum dubium has also been mistaken for Anoectangium (e.g., W. Bell 639, CHR 566204), a confusion definitively resolved by the difference in laminal cell KOH reaction (yellow in Anoectangium vs red in Bryoerythrophyllum). Zander (1993) noted the similarity of Syntrichia abruptinervis to Anoectangium species, and listed distinguishing features, including KOH reaction (red in Syntrichia). Confusion by H.N. Dixon of the type of Anoectangium bellii with Ardeuma recurvirostrum (as Hymenostylium recurvirostre) is discussed under the latter species.
NI: S. Auckland (Maungapōhatu), Wellington (Ruapehu); SI: Nelson, Marlborough, Canterbury, Westland, Otago; Ch. Recorded from A by Vitt (1979).
Nearly cosmopolitan, including mainland Australia (Stone 1990), but not known from southern South America or southern Africa.
An uncommon species, usually found in sheltered positions on damp rock, including schist, granite, and greywacke. Associated mosses include Austrohondaella limata, Bryoerythrophyllum recurvirostrum, and Tridontium tasmanicum.
The species is recorded from c. 1000–2000 m elevation.
A. aestivum is widely distributed in the South I. (principally on the wetter, western side). Very few collections are known from the North I.
This moss was first recorded for N.Z. by Hooker (1867) under the name "A. compactum Schwaegr.", a species already known from Europe. Dixon (1926) described the N.Z. species as A. bellii, differing from the European plant in having "foliis multo angustioribus, linearibus vel lineari-lanceolatis, peracutis, et nervo dorsa minute distincte scaberulo." He continued, "The New Zealand species is much like the European A. compactum [A. aestivum], but differs clearly in the narrower, very acute leaves, with a sharp point mostly formed by the excurrent or percurrent nerve, which is finely but distinctly scaberulous at back." However, the costal surface ornamentation and leaf shape of A. bellii fall within the range of northern hemisphere material, including (for leaf shape) that illustrated in Hedwig’s protologue (Hedwig 1801, Tab. II. f. 6). The N.Z. taxon A. bellii is here accepted as a synonym with the widespread and very variable A. aestivum, as proposed by Sollman (2023).
The plethora of isotype specimens for Anoectangium bellii is due to confusion attributable to H.N. Dixon, and explained in the notes for Ardeuma recurvirostrum elsewhere in this Flora.
A second species of Anoectangium, A. stracheyanum, has been recorded for N.Z. (van Zanten & Sollman 2023). The species is widely distributed in both hemispheres and seemingly often in mountainous regions (van Zanten & Sollman 2023). The N.Z. record is based on specimens from several alpine and subalpine South I. localities (A.J. Fife 6468, CHR 405608; A.J. Fife 5947, CHR 104939; A.J. Fife 7391, CHR 406531). The species is not considered further in this treatment.